Sustainable Carbon Source Utilization for Indole-3-Acetic Acid Production by Marine Pseudomonas fluorescens BCPBMS-1 Using the One-Factor-at-a-Time Approach
| Received 01 Nov, 2024 |
Accepted 24 Dec, 2024 |
Published 30 Jun, 2025 |
Background and Objective: Indole-3 acetic acid (IAA) is a phytohormone that stimulates plant growth and facilitates plant development. Indole-3-acetic acid is a widely occurring endogenous plant hormone that has been shown to enhance root development and morphology. The current study involved the extraction of IAA from marine Pseudomonas fluorescensBCPBMS-1. Materials and Methods: Indole acetic acid (IAA) production in P. fluorescens was screened using Luria broth with L-tryptophan and detected calorimetrically with the Salkowski reagent. Optimization involved testing various factors, including temperature, pH, salinity, incubation time and different carbon and nitrogen sources. The IAA concentration was measured spectrophotometrically at 530 nm using a standard IAA curve. Results: The optimized conditions for maximum IAA production are 30°C, pH 6, 10 ppt salinity and 72 hrs of incubation. Glucose and yeast extract were the most effective carbon and nitrogen sources, respectively, while soybean husk was the best low-cost alternative. Mass-scale synthesis under these conditions yielded 0.72 μg/mL IAA using cost-effective substrates and precise parameters. Conclusion: Microbes’ ability to produce indole-3-acetic acid (IAA) offers an economical replacement for expensive and unreliable chemical IAA synthesis methods. According to this study, soybean husk represents a more economical carbon source for the production of IAA.
| Copyright © 2025 Venkatraman et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Over the past few decades, there has been a consistent increase in the world population and the subsequent need for food. As a result, there is a pressing need for advancements in agricultural production. Indole-3 acetic acid (IAA) is a plant hormone that is essential for the growth and development of plants. Indole-3-acetic acid (IAA) is the predominant natural auxin in plants and is known to enhance root growth and shape. This is thought to improve nutrient uptake from the soil1. Numerous researchers have paid close attention to the role of IAA in the intricate interplay between the microflora in the rhizosphere and the host plant, which is based on a continuous exchange of substances and signals2-5. The synthesis of elevated quantities of IAA by rhizosphere bacteria has been proven to inhibit rather than promote root growth6,7. This is in contrast to the production of IAA by microorganisms commonly found in the rhizosphere of plants, such as Pseudomonas spp. and Rhizobium spp. which are frequently linked to their potential to stimulate plant growth8-11.
Plants and soil microbes in the rhizosphere can interact in ways that are either positive, neutral, variable, or detrimental to plant growth. According to Cartwright et al.12, these organisms’ beneficial activities include fixing nitrogen, producing siderophores, chitinase, antibiotics and dissolving phosphate and other nutrients, as well as producing or altering the concentrations of plant hormones such as IAA, gibberellic acid, cytokinin and ethylene. Gram-negative, aerobic, polarly flagellated rods are the organisms that belong to the Pseudomonas genus. They are known to grow on basic medium and are aggressive invaders of the rhizosphere of a variety of crop plants13,14.
Marine Pseudomonas spp. is a member of the microorganism community in marine extreme environments15 and represents untapped novel bioactive substances16. Marine isolates of Pseudomonas spp. are found in diverse ecosystems, including coastal regions, the deep sea and more extreme environments17.
Few studies have been undertaken on optimizing marine microbial IAA production, leaving a knowledge gap. Therefore, it is essential to optimize the production medium for IAA from marine Pseudomonas fluorescens BCPBMS-1. This technique is the most straightforward to use and greatly aids in the identification of the essential variables that influence IAA production. This study investigated the sustainable utilization of carbon sources for the production of indole-3-acetic acid (IAA) by marine Pseudomonas fluorescens BCPBMS-1 using the one-factor-at-a-time approach.
MATERIALS AND METHODS
Study area: Sponges have been collected from the Gulf of Mannar, India’s Southeast Coast (Latitude 9°5’N, Longitude 79°5’E). Shortly after collection, the sponge sample was transferred to a sterile plastic bag and transported frozen to the laboratory for the isolation of bacteria. The samples were collected and study performed in April, 2010.
Screening for indole acetic acid (IAA) production: Screening was done by using Luria broth (HI-Media) supplemented with L-tryptophan. A loop full of P. fluorescens was inoculated in 10 mL of Luria broth supplemented with 0.1% of L-tryptophan and incubated for 72 hrs at 30°C. Then, the culture was centrifuged at 10,000Xg for 10 min. and the supernatant was collected. The 1 mL of supernatant was allowed to react with 2 mL of Salkowski reagent (1 mL of 0.5 M FeCl3 in 50 mL of 35% HClO4) at 30°C for 30 min. Pink color development indicated the presence of IAA4.
Medium optimization in shake-flask cultivation
Experimental design for optimizing IAA production using low-cost substrates by one factor at a time method: The effects of various factors on the production of IAA were tested individually. These factors included temperature (ranging from 20-35°C with a 5°C interval), pH (ranging from 5-8 with a 1-unit interval), salinity (ranging from 2-12 ppt with a 2 ppt interval), the incubation period (ranging from 24-96 hrs with a 24 hrs interval) and different carbon and nitrogen sources. The carbon sources tested were 3% glucose, sucrose and fructose, while the nitrogen sources tested were yeast extract, peptone and beef extract at a concentration of 0.5%. Additionally, economically cheaper sources such as soybean husk, wheat bran and molasses were tested at a concentration of 3%. The experiments were conducted under shaken conditions at 150 rpm.
Estimation of IAA: The P. fluorescens culture was centrifuged at 13000Xg for 10 min and the supernatant was collected. The presence of IAA was measured in a spectrophotometer by adding 2 mL of Salkowski reagent to 1 mL of supernatant and incubating for 30 min. The optical density was read at 530 nm. The recorded OD values were plotted in a standard curve prepared from commercially available IAA and their concentration was calculated18.
RESULTS AND DISCUSSION
Screening for indole acetic acid (IAA) production: The initial development of roots, either through the elongation of primary roots or the multiplication of lateral and adventitious roots, is beneficial for young seedlings. This enables them to firmly anchor themselves to the soil and acquire water and nutrients from their surroundings, thereby improving their chances of survival. Root colonization by bacteria known as “plant growth promoting” bacteria (PGPR) can boost plant growth19-22. Microbial synthesis of the plant-growth regulator IAA23,24 can result in to improved plant growth.
In the present study, P. fluorescens which was isolated from the Callyspongia diffusa produced indole acetic acid. It was screened by using Luria broth supplemented with L-tryptophan and the addition of the Salkowski reagent. A positive result was indicated by pink color. Bhadbhade et al.11 reported that a higher level of IAA production was produced by Pseudomonas spp. Shokri and Emtiazi25 also reported that indole acetic acid was produced higher level in Pseudomonas spp., compared to Azotobacter spp.
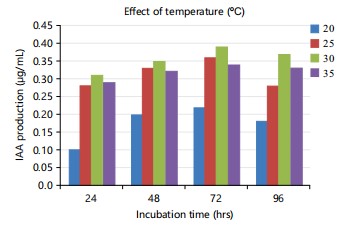
Optimization by one factor at a time: The results showed that the highest IAA production of 0.39 μg/mL occurred at a temperature of 30°C, indicating that this temperature was the most beneficial for IAA production (Fig. 1). According to Shokri and Emtiazi25, the ideal temperature for producing IAA was found to be 30°C. A study conducted by Aldesuquy et al.26 revealed that temperatures between 25-30°C were optimal for both the development and synthesis of indole-3-acetic acid (IAA) in Streptomyces spp. According to Apine and Jadhav27, a temperature of 30°C was found to be optimal for the synthesis of IAA by the Pantoea agglomerans strain PVM.
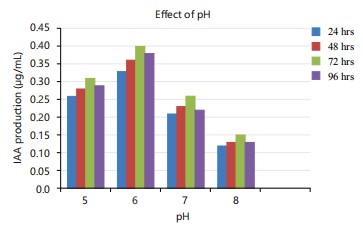
In this study, different pH levels were used to see how they impacted the production of IAA. At pH 6, the most was made (0.4 μg/mL) and at pH 8, the least was made (0.12 μg/mL) (Fig. 2). According to Huddedar et al.28 Acinetobacter baumannii A16 and Acinetobacter A18 strains can make the most IAA when the pH is 7.0. A study by Aldesuquy et al.26 observed that highest amount of IAA was produced when the pH was 7.0. Mandal et al.29 reported that the highest level of IAA production came at pH 7.2. According to Yurekli et al.30 the production of IAA is suited to slightly acidic conditions with a pH of 7.5 or higher.
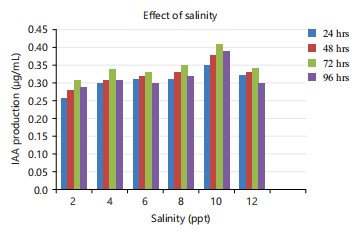
In the present observation, 10 ppt salinity showed the maximum production of 0.41 μg mL and at 2 ppt salinity only 0.26 μg/mL IAA was produced (Fig. 3). Ravikumar et al.31 reported that A. brasiliense isolated from mangrove roots produced the highest level of IAA at a concentration of 1% NaCl. According to Sachdev et al.32, the optimal NaCl concentration for IAA production in Klebsiella spp., is 0.5 w/v.
Regarding incubation time, maximum production was observed at 72 hrs and the minimum was observed at 24 hrs (Fig. 1-3). Castillo et al.33 observed IAA accumulation in Azospirillum and Arthrobacter spp., during the stationary phase. The current investigation supported this finding as well. Chung and Tzeng34 found that fungi U. maydis produced IAA after 3 days of incubation.
Shokri and Emtiazi25 similarly found that 72 hrs was the best incubation time for IAA production. Lin and Xu35 observed the same thing in Streptomyces spp. The decrease in IAA synthesis after 72 hrs could be attributed to the release of IAA-degrading enzymes such as IAA oxidase and peroxidase, as previously observed in Rhizobium spp., isolated from Cajanus cajan36,37.
|
|
|
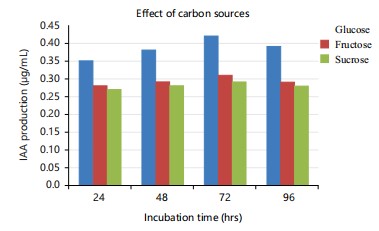
Glucose exhibited the highest production (0.42 μg/mL) among the carbon sources utilized, as seen in Fig. 4. The highest production of IAA was reported while using a 3% glucose concentration, as determined using a one-factor-at-a-time approach in the current study. The high IAA synthesis in the glucose-containing medium may be attributed to the more efficient use of this easily utilizable glucose in comparison to other sources38. Rhizobium spp. derived from Cajanus cajan similarly exhibited the highest production of indole-3-acetic acid (IAA) in a medium containing glucose, as previously documented by Datta and Basu36. In their study, Narayana et al.38 found that the highest generation of IAA occurred when 1% glucose was employed as the carbon source, using a one-factor-at-a-time method.
|
|
|
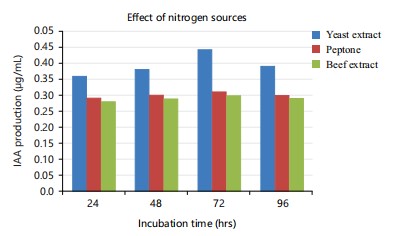
Among the nitrogen sources tested, yeast extract showed the maximum production of IAA (0.47 μg/mL) and minimum production was observed with beef extract (0.28 μg/mL) (Fig. 5). Narayana et al.38also observed the highest IAA production from Streptomyces albidoflavus in the medium containing yeast extract as a nitrogen source.
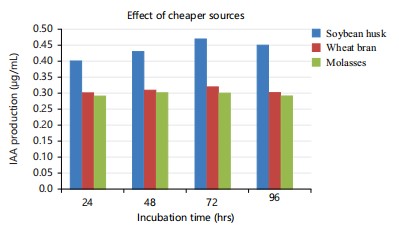
Out of the cheaper sources that were evaluated, the highest level of production was reported with soybean husk, which had a concentration of 0.48 μg/mL. The lowest concentration was recorded with molasses at 0.29 μg/mL (Fig. 6). In this investigation, it was determined that soybean husk is the most cost-effective source. Debebe et al.39 employed soybean supplement to stimulate the production of indole-3-acetic acid (IAA) by Pseudomonas spp., Rubio et al.40 found that Azotobacter spp., Enterobacter spp. and Pseudomonas spp. exhibited a preference for soy flour as the substrate for indole-3-acetic acid (IAA) production.
Mass production of IAA: A concentration of 0.72 μg/mL was achieved during large-scale production using the optimized physical as well as chemical parameters and a most cost-effective carbon source (soya bean husk). Nenwani et al.41 documented that the concentration of indole-3-acetic acid (IAA) produced was measured to be 11.45 μg/mL. According to Khakipour et al.42 the P. fluorescens strains produced indole-3-acetic acid (IAA) in quantities ranging from 0 to 31.6 mg/L, while P. putida produced IAA in quantities ranging from 0 to 24.08 mg/L. According to Leinhos and Vacek43 P. fluorescens produced 1.6-3.3 mg/L of IAA, but Prikryl et al.44 detected auxin production ranging from 0.01-3.93 mg/L. Wu et al.45 reported the observation of a significant generation of indole-3-acetic acid (IAA) by genetically modified E.coli, reaching a high concentration of 7 g/L. Nevertheless, to sustain a high level of IAA synthesis, this genetically modified bacterium relied on the presence of an inducer and selective pressure. The technique may not be suited for large-scale industrial production of IAA due to the high expense necessary to maintain stability in bacterial IAA production.
CONCLUSION
This study improved IAA production by marine Pseudomonas fluorescens BCPBMS-1 using a one-factor-at-a-time method to investigate formulations that are most suited to economically viable carbon sources for commercial production of IAA. A suitable medium for better IAA production was successfully established and IAA production was increased by adjusting the culture parameters using one factor at a time method. The findings suggest that the most cost-effective carbon source for IAA production is soya bean husk.
SIGNIFICANCE STATEMENT
This study highlights the potential of Pseudomonas fluorescens BCPBMS-1 for cost-effective, large-scale production of indole-3-acetic acid (IAA), providing a sustainable alternative to chemical synthesis and conventional microbial processes. The optimization of production parameters establishes a scalable bioprocess framework, promoting environmentally friendly solutions to reduce dependence on harmful agricultural chemicals and enhance sustainable agricultural practices.
REFERENCES
- Vessey, J.K., 2003. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil, 255: 571-586.
- Saeed, Q., W. Xiukang, F.U. Haider, J. Kučerik and M.Z. Mumtaz et al., 2021. Rhizosphere bacteria in plant growth promotion, biocontrol, and bioremediation of contaminated sites: A comprehensive review of effects and mechanisms. Int. J. Mol. Sci., 22.
- Costacurta, A. and J. Vanderleyden, 1995. Synthesis of phytohormones by plant-associated bacteria. Crit. Rev. Microbiol., 21: 1-18.
- Patten, C.L. and B.R. Glick, 1996. Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol., 42: 207-220.
- Persello-Cartieaux, F., L. Nussaume and C. Robaglia, 2003. Tales from the underground: Molecular plant-rhizobacteria interactions. Plant Cell Environ., 26: 189-199.
- Xie, H., J.J. Pasternak and B.R. Glick, 1996. Isolation and characterization of mutants of the plant growth-promoting rhizobacterium Pseudomonas putida GR12-2 that overproduce indoleacetic acid. Curr. Microbiol., 32: 67-71.
- Ratnaningsih, H.R., Z. Noviana, T.K. Dewi, S. Loekito, S. Wiyono, Abdul Gafur and S. Antonius, 2023. IAA and ACC deaminase producing-bacteria isolated from the rhizosphere of pineapple plants grown under different abiotic and biotic stresses. Heliyon, 9.
- Antoun, H., C.J. Beauchamp, N. Goussard, R. Chabot and R. Lalande, 1998. Potential of Rhizobium and Bradyrhizobium Species as Plant Growth Promoting Rhizobacteria on Non-Legumes: Effect on Radishes (Raphanus sativus L.). In: Molecular Microbial Ecology of the Soil: Results from an FAO/IAEA Co-Ordinated Research Programme, 1992–1996, Hardarson, G. and W.J. Broughton (Eds.), Springer, Dordrecht, Netherlands, ISBN: 978-94-017-2321-3, pp: 57-67.
- Barazani, O. and J. Friedman, 1999. Is IAA the major root growth factor secreted from plant-growth-mediating bacteria? J. Chem. Ecol., 25: 2397-2406.
- Patten, C.L. and B.R. Glick, 2002. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol., 68: 3795-3801.
- Bhadbhade, B.J., S.S. Sarnaik and P.P. Kanekar, 2002. Biomineralization of an organophosphorus pesticide, Monocrotophos, by soil bacteria. J. Appl. Microbiol., 93: 224-234.
- Cartwright, D.K., W.S. Chilton and D.M. Benson, 1995. Pyrrolnitrin and phenazine production by Pseudomonas cepacia, strain 5.5B, a biocontrol agent of Rhizoctonia solani. Appl. Microbiol. Biotechnol., 43: 211-216.
- Blanco-Romero, E., D. Durán, D. Garrido-Sanz, M. Redondo-Nieto, M. Martín and R. Rivilla, 2023. Adaption of Pseudomonas ogarae F113 to the rhizosphere environment-the AmrZ-FleQ hub. Microorganisms, 11.
- Kaneko, H., H. Takami, A. Inoue and K. Horikoshi, 2000. Effects of hydrostatic pressure and temperature on growth and lipid composition of the inner membrane of barotolerant pseudomonas sp. BT1 isolated from the deep-sea. Biosci. Biotechnol. Biochem., 64: 72-79.
- Isnansetyo, A. and Y. Kamei, 2009. Bioactive substances produced by marine isolates of Pseudomonas. J. Ind. Microbiol. Biotechnol., 36: 1239-1248.
- Engel, S., P.R. Jensen and W. Fenical, 2002. Chemical ecology of marine microbial defense. J. Chem. Ecol., 28: 1971-1985.
- Khan, Z. and S.L. Doty, 2009. Characterization of bacterial endophytes of sweet potato plants. Plant Soil, 322: 197-207.
- Kloepper, J.W., J. Leong, M. Teintze and M.N. Schroth, 1980. Enhanced plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature, 286: 885-886.
- Saleh, S.S. and B.R. Glick, 2001. Involvement of gacS and rpoS in enhancement of the plant growth-promoting capabilities of Enterobacter cloacae CAL2 and UW4. Can. J. Microbiol., 47: 698-705.
- Ryu, C.M., M.A. Farag, C.H. Hu, M.S. Reddy, H.X. Wei, P.W. Pare and J.W. Kloepper, 2003. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A., 100: 4927-4932.
- Ryu, C.M., M.A. Farag, C.H. Hu, M.S. Reddy, J.W. Kloepper and P.W. Paré, 2004. Bacterial volatiles induce systemic resistance in Arabidopsis. Plant Physiol., 134: 1017-1026.
- Lambrecht, M., Y. Okon, A.V. Broek and J. Vanderleyden, 2000. Indole-3-acetic acid: A reciprocal signalling molecule in bacteria-plant interactions. Trends Microbiol., 8: 298-300.
- Bano, N. and J. Musarrat, 2004. Characterization of a novel carbofuran degrading Pseudomonas sp. with collateral biocontrol and plant growth promoting potential. FEMS Microbiol. Lett., 231: 13-17.
- Lebrazi, S., M. Fadil, M. Chraibi and K. Fikri-Benbrahim, 2020. Screening and optimization of indole-3-acetic acid production by Rhizobium sp. strain using response surface methodology. J. Genet. Eng. Biotechnol., 18.
- Shokri, D. and G. Emtiazi, 2010. Indole-3-acetic acid (IAA) production in symbiotic and non-symbiotic nitrogen-fixing bacteria and its optimization by Taguchi design. Curr. Microbiol., 61: 217-225.
- Aldesuquy, H.S., F.A. Mansour and S.A. Abo-Hamed, 1998. Effect of the culture filtrates of Streptomyces on growth and productivity of wheat plants. Folia Microbiol., 43: 465-470. .
- Apine, O.A. and J.P. Jadhav, 2011. Optimization of medium for indole-3-acetic acid production using Pantoea agglomerans strain PVM. J. Appl. Microbiol., 110: 1235-1244.
- Huddedar, S.B., A.M. Shete, J.N. Tilekar, S.D. Gore, D.D. Dhavale and B.A. Chopade, 2002. Isolation, characterization and plasmid pUPI126-mediated indole-3-acetic acid production in Acinetobacter strains from rhizosphere of wheat. Appl. Biochem. Biotechnol., 102: 21-39.
- Mandal, S., M. Mandal, A. Das, B. Pati and A. Ghosh, 2009. Stimulation of indoleacetic acid production in a Rhizobium isolate of Vigna mungo by root nodule phenolic acids. Arch. Microbiol., 191: 389-393.
- Yurekli, F., H. Geckil and F. Topcuogul, 2003. The synthesis of indole-3-acetic acid by the industrially important white-rot fungus Lentinus sajor-caju under different culture conditions. Mycol. Res., 107: 305-309.
- Ravikumar, S., K. Kathiresan, S.T.M. Ignatiammal, M.B. Selvam and S. Shanthy, 2004. Nitrogen-fixing azotobacters from mangrove habitat and their utility as marine biofertilizers. J. Exp. Mar. Biol. Ecol., 312: 5-17.
- Sachdev, D.P., H.G. Chaudhari, V.M. Kasture, D.D. Dhavale and B.A. Chopade, 2009. Isolation and characterization of indole acetic acid (IAA) producing Klebsiella pneumoniae strains from rhizosphere of wheat (Triticum aestivum) and their effect on plant growth. Indian J. Exp. Biol., 47: 993-1000.
- Castillo, P., R. Molina, A. Andrade, A. Vigliocco, S. Alemano and F.D. Cassán, 2015. Phytohormones and Other Plant Growth Regulators Produced by PGPR: The Genus Azospirillum. In: Handbook for Azospirillum: Technical Issues and Protocols, Castillo, P., R. Molina, A. Andrade, A. Vigliocco, S. Alemano and F.D. Cassán (Eds.), Springer, Cham, Switzerland, ISBN: 978-3-319-06542-7, pp: 115-138.
- Chung, K.R. and D.D. Tzeng, 2004. Biosynthesis of indole-3-acetic acid by the gall-inducing fungus Ustilago esculenta. J. Biol. Sci., 4: 744-750.
- Lin, L. and X. Xu, 2013. Indole-3-acetic acid production by endophytic Streptomyces sp. En-1 isolated from medicinal plants. Curr. Microbiol., 67: 209-217.
- Datta, C. and P.S. Basu, 2000. Indole acetic acid production by a Rhizobium species from root nodules of a leguminous shrub, Cajanus cajan. Microbiol. Res., 155: 123-127.
- Sridevi, M. and K.V. Mallaiah, 2007. Production of indole-3-acetic acid by Rhizobium isolates from Sesbania species. Afr. J. Microbiol. Res., 1: 125-128.
- Narayana, J.K.P., P. Prabhakar, M. Vijayalakshmi, Y. Venkateswarlu and P.S.J. Krishna, 2007. Biological activity of phenylpropionic acid isolated from a terrestrial Streptomycetes. Pol. J. Microbiol., 56: 191-197.
- Lata, D.L., O. Abdie and Y. Rezene, 2024. IAA-producing bacteria from the rhizosphere of chickpea (Cicer arietinum L.): Isolation, characterization, and their effects on plant growth performance. Heliyon, 10.
- Torres-Rubio, M.G., S.A. Valencia-Plata, J. Bernal Castillo and P. Martinez-Nieto, 2000. Isolation of enterobacteria, Azotobacter sp. and Pseudomonas sp., producers of indole-3-acetic acid and siderophores, from colombian rice rhizosphere. Rev. Latinoam. Microbiolia, 42: 171-176.
- Nenwani, V., P. Doshi, T. Saha and S. Rajkumar, 2010. Isolation and characterization of a fungal isolate for phosphate solubilization and plant growth promoting activity. J. Yeast Fungal Res., 1: 9-14.
- Khakipour, N., K. Khavazi, H. Mojallali, E. Pazira and H. Asadirahmani, 2008. Production of auxin hormone by fluorescent pseudomonads. Am. Eurasian J. Agric. Environ. Sci., 4: 687-692.
- Leinho, V. and O. Vacek, 1994. Biosynthesis of auxins by phosphate solubilizing rhizobacteria from wheat (Triticum aestivum) and rye (Secale cereale). Microbiol. Res., 149: 31-35.
- Přikryl, Z., V. Vančura and M. Wurst, 1985. Auxin formation by rhizosphere bacteria as a factor of root growth. Biol. Plant., 27: 159-163.
- Wu, H., J. Yang, P. Shen, Q. Li and W. Wu et al., 2021. High-level production of indole-3-acetic acid in the metabolically engineered Escherichia coli. J. Agric. Food Chem., 69: 1916-1924.
How to Cite this paper?
APA-7 Style
Venkatraman,
V., Ramasamy,
L., Singaram,
J.l. (2025). Sustainable Carbon Source Utilization for Indole-3-Acetic Acid Production by Marine Pseudomonas fluorescens BCPBMS-1 Using the One-Factor-at-a-Time Approach. Asian Journal of Biological Sciences, 18(2), 300-307. https://doi.org/10.3923/ajbs.2025.300.307
ACS Style
Venkatraman,
V.; Ramasamy,
L.; Singaram,
J.l. Sustainable Carbon Source Utilization for Indole-3-Acetic Acid Production by Marine Pseudomonas fluorescens BCPBMS-1 Using the One-Factor-at-a-Time Approach. Asian J. Biol. Sci 2025, 18, 300-307. https://doi.org/10.3923/ajbs.2025.300.307
AMA Style
Venkatraman
V, Ramasamy
L, Singaram
Jl. Sustainable Carbon Source Utilization for Indole-3-Acetic Acid Production by Marine Pseudomonas fluorescens BCPBMS-1 Using the One-Factor-at-a-Time Approach. Asian Journal of Biological Sciences. 2025; 18(2): 300-307. https://doi.org/10.3923/ajbs.2025.300.307
Chicago/Turabian Style
Venkatraman, Vasanthabharathi, Lakshminarayanan Ramasamy, and Jaya lakshmi Singaram.
2025. "Sustainable Carbon Source Utilization for Indole-3-Acetic Acid Production by Marine Pseudomonas fluorescens BCPBMS-1 Using the One-Factor-at-a-Time Approach" Asian Journal of Biological Sciences 18, no. 2: 300-307. https://doi.org/10.3923/ajbs.2025.300.307

This work is licensed under a Creative Commons Attribution 4.0 International License.