Invasive Chromolaena odorata Infestation Supports Increased Arthropod Diversity in Southern Nigeria
-
Jane I. Otabor

Department of Biological Sciences, Federal University Wukari, PMB 1020, Taraba State, Nigeria
Mildred O. UdemaduDepartment of Animal and Environmental Biology, University of Benin, PMB 1154, Benin, Nigeria
Nosa OsaweDepartment of Animal and Environmental Biology, University of Benin, PMB 1154, Benin, Nigeria
Cornell O. IsikhuemenDepartment of Animal and Environmental Biology, University of Benin, PMB 1154, Benin, Nigeria
May N. OkaforDepartment of Fine and Applied Arts, University of Nigeria, Nsukka, Nigeria
Osariyekemwen UyiDepartment of Animal and Environmental Biology, University of Benin, PMB 1154, Benin, Nigeria
| Received 14 Nov, 2024 |
Accepted 15 Jan, 2025 |
Published 31 Mar, 2025 |
Background and Objective: The highly invasive Chromolaena odorata (Siam weed) is well recognized as a plant that alters ecosystem composition and trophic functionality. Although these changes are mostly deleterious to native plant species, little is known about their impact on associated arthropod communities. This study investigated how C. odorata invasion influences arthropod community assemblage. This study compared the diversity, richness and evenness of arthropods in sites without C. odorata (0% C. odorata-infested site) and with C. odorata (>50% C. odorata-infested sites). Materials and Methods: Pitfall traps and beating tray sampling techniques were used to sample both epigeal and arboreal arthropods, respectively and collected arthropods were identified at the family level. Diversity at the family level was analyzed using richness, Shannon and Simpson indices, calculated in PAST software (version 4.03). Results: The results suggest that sites with >50% C. odorata coverage supported higher arthropod family richness, diversity and evenness compared to the 0% C. odorata uninfested site, contradicting the initial expectations. Although a 51% Jaccard similarity index suggests a moderate overlap in family composition within the two study sites, this study contrasts the typical expectation of reduced arthropod diversity following the invasion of an aggressive plant species. Conclusion: The study uniquely demonstrates that C. odorata invasion may enhance certain aspects of arthropod community structure, providing new insights into the complexity of ecosystem responses to invasive species.
| Copyright © 2025 Otabor et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Chromolaena odorata, commonly known as Siam weed or bitter bush, is an herbaceous to woody perennial shrub belonging to the family Asteraceae (the sunflower family) and is native to North America1,2. This plant typically reaches heights of 1-3 m and produces dense clusters of small, pale blue to purple flowers3. When crushed, its leaves release a strong odor, thought to deter herbivores and enhance their resilience in various environments4. This plant has gained attention for its medicinal properties, being widely used to treat wounds, burns and skin infections. It has demonstrated anticancer, antidiabetic, antinociceptive, anti-hepatotoxic, anti-inflammatory, antimicrobial and antioxidant effects5. However, C. odorata is often regarded as a weed and a matter of global concern, especially in exotic regions.
The earliest recorded case of C. odorata as an invasive species dates back to 1872 in India, followed by its spread to Sumatra, Indonesia, in 1934. Today, C. odorata is widespread across numerous countries in Asia, Western and Southern Africa and Australia1,6. In South Africa, it has been regarded as the second most prevalent invasive plant species and is considered one of the world’s most destructive weed invaders7,8. Following its introduction from Sri Lanka into Southern Nigeria in 19379, it has reached critical levels in Nigeria10-12, Cameroon, Ghana and other African regions13 and is now recognized as one of the worst weeds in Nigeria and West Africa. The plant was first documented in Nigeria near Enugu State in 194210 and has since rapidly expanded throughout Southern Nigeria, particularly in areas with high rainfall and humidity14-16. The radical spread of C. odorata in Nigeria can be attributed to several factors: Its exceptional reproductive capacity, adaptability to the local climate, ability to alter soil properties in ways that render the soil unsuitable for other competing species and its tendency to form dense thickets that out-compete native plants, leading to reduced abundance or even elimination of indigenous species12,17,18. Such large-scale plant invasion contributes to ecological homogenization, disrupting food chains that support higher trophic levels18.
The proliferation of invasive alien weeds remains a paramount concern for preserving natural habitats and their resident species. Invasive plants, like C. odorata, sometimes unpredictably impact ecosystems in multiple ways, such as increasing soil nitrogen, altering soil eco-hydrological conditions, trapped sediment composition and microclimatic factors8,18. By forming dense populations, these invaders outcompete native plant species, reducing biodiversity through direct competition and disrupting multiple trophic levels within ecological communities1,19-23. Limited attention has been devoted to understanding the impact of C. odorata on community-level dynamics of higher trophic feeders in Nigeria, with few studies exploring these interactions. For instance, Mgobozi et al.24 studied the effects of C. odorata on spider diversity, Rizali et al.25 examined how ants respond to C. odorata and Buchori et al.26 documented the local arthropod fauna associated with this invasive plant. However, studies within the study area have yet to investigate the response of arthropods to the presence of the invasive plant.
Arthropods are particularly valuable for studying community-level responses to plant invasions. As a diverse and ecologically significant taxonomic group with a global presence, they respond quickly to environmental changes, making them effective indicators of both local and broader landscape changes. Plant invasions can alter arthropod composition by modifying resource availability, affecting both herbivores that feed on the plants and the higher trophic levels that depend on these herbivores as prey27-30. A change in the resource-availability dynamics may force shifts in the community composition of associated arthropods through three major mechanisms: (1) Selection, favoring species best adapted to the remaining resources, (2) Migration or dispersal of species to other areas and (3) Eventual speciation as some species evolve over the long term to adapt to the altered conditions. Selection occurs as some arthropods may be able to survive and reproduce within this altered landscape, benefiting from the changes introduced by the invasive plant. In resource scarcity, they may die out or take the alternative: Dispersal. Arthropods may tend to migrate outside their zones of resthece unavailability, typically to more resource-available regions. Furthermore, changes in the plant composition change the landscape, which, over time, may evolutionarily drive the formation of new species.
Although C. odorata has been in Nigeria for eight decades, no scientific evidence yet indicates that it has driven speciation among arthropods. Any small evolutionary changes may be too subtle to produce noticeable taxonomic divergence. Since arthropods can be efficiently sampled and have relatively short lifespans, they offer a practical way to observe responses to C. odorata invasion, allowing one to detect ecological changes and species turnover within a relatively short time frame. Current studies hypothesize that C. odorata invasion will result in lower arthropod diversity, evenness and overall community homogenization. To test this, the current study selected rural areas in Edo State, Southern Nigeria, where C. odorata has been well-established. In these minimally disturbed areas with low anthropogenic activity, the current study will assess and compare arthropod abundance and diversity between C. odorata-infested and un-infested sites. Specifically, the current study will evaluate the family richness, evenness and diversity of foliage-dwelling and ground-active arthropods across invaded and non-invaded sites.
MATERIALS AND METHODS
Study area and sample site: The study was conducted from October to November 2023 in Ogua community, Ovia North East Local Government Area, Edo State, Nigeria. One large fallow land (GPS: Latitude 6°31'30"N and Longitude 5°32'25"E) with ~0% C. odorata infestation and another (GPS: Latitude 6°31'27"N and Longitude 5°32'26"E) with 50% C. odorata were investigated in this study. To ensure independence in the arthropod community, each site was separated by at least 500 m, as suggested by Effah et al.21. For better reference, the site with 0% C. odorata will subsequently be referred to as site 1 and that with >50% C. odorata, site 2.
Site 1 is a forested area with huge trees, crawling plants, shrubs, grasses and floor litter and measures about 100×100 m2. It consisted of a partial canopy cover and was dominated by Cyphostemma adenocaule (Vitaceae) and Palisota hirsuta (Commelinaceae).
The dominant vegetation at site 2 were Chromolaena odorata, Croton hirtus (Euphorbiaceae), Myrianthus arboreus (Urticaceae), Setaria barbata (Poaceae) and Centrosema pubescens (Fabaceae).
Arthropod sampling: Two sampling techniques, pitfall trapping and beating-tray, were used in the arthropod collection. Afterward, all arthropods collected were stored using paper envelopes or vials containing 70% ethanol where appropriate. All collected arthropods were sorted into morpho-species using morphological differences31. Subsequently, they were identified as part of the taxa group (Order, Family) using the help of standard insect guides32.
Pitfall trapping: At each site, 15 transparent pitfall traps (75 mL plastic cups, 8 cm diameter and 9 cm depth) were installed on the ground and flushed to the surface. Each trap was placed 2 m apart. Traps were covered for 5 days before sampling began to eliminate the digging-in effect33, after which about 25 mL of water containing a few drops of liquid soap to break surface tension was added to each trap. Trapped arthropods were collected three times (i.e., at 24, 48, 72 hrs after the opening of the trap).
Beating tray technique: A white kite-like tray measuring about 1 m2 was placed below randomly selected C. odorata plants in the >50% C. odorata region and gently beaten with a beating stick continuously for about 5 min. Quick collection of all arthropods on the beating tray was done using an aspirator and put in a killing jar. The same was done on random plants on the 0% C. odorata plots. Ten collections were made for each site, making a total of twenty collections. The insects were collected between 8:00-10:00 hrs.
Data analysis: The arthropod families were organized and tabulated in Microsoft Excel. For diversity analysis at the family level, richness, Shannon and Simpson indices were calculated using PAST software version 4.0334. The Shannon diversity index, calculated in PAST35, measures diversity on a scale from 0, where one family dominates, to higher values indicating a wider variety of families with a more balanced representation. To estimate uniformity in family distribution, the current study calculated the evenness index, standardizing Shannon’s measure from 0-1, with values close to 1 indicating equal representation across families. Lower evenness values reflect uneven representation, while higher values suggest more balanced distributions. As an inverse measure, dominance highlights species dominance within the community, where higher dominance values indicate less even distribution across species. Also, the current study plotted a rarefaction curve that represents the cumulative family richness with increased sampling coverage and sampling effort. Finally, the current study calculated the Jaccard index to assess the compositional similarity between C. odorata-infested and non-infested areas.
RESULTS
A total of 488 arthropods belonging to 54 families were sampled from both sites (0 and 50% C. odorata). Site 1 (0% C. odorata infestation) had 34 families, while 42 families were found in the infested site (Table 1). The most abundant family was Formicidae, with 183 individuals (37.5%), followed by the family Evaniidae with 36 individuals (7.4%). Family Scarabaeidae and Muscidae both had 24 individuals each (4.9%) and every other family had a relative abundance of less than 5%. With pitfall and beating tray sampling methods being the only sampling methods used, 346 individuals belonging to 38 families were captured using the pitfall technique, while the beating tray technique sampled 142 individuals belonging to 28 families from both sites. From site 1 (0% C. odorata), 202 individuals were recorded and 286 from site 2 (> 50% C. odorata). From site 1 (0% C. odorata), 202 individuals were recorded and 286 from site 2 (>50% C. odorata). The most abundant family in site 1 and 2 was the Formicidae family, with 102 and 81 individuals in site 1 and 2, respectively (Table 1). Finally, the Jaccard index was 0.511, indicating that around 51.1% of the arthropod families are shared between the two sites.
Diversities of foliage and ground-dwelling arthropods are shown in Table 2. Family richness was generally higher in the infested site (site 2) across both sampling methods, with 17 families recorded from foliage samples in site 2 compared to 15 in site 1 and 32 families from ground samples in site 2 compared to 24 in site 1. In terms of individual arthropods, ground arthropod abundance was higher in the infested site compared to the uninfested site, while foliage arthropods demonstrated the opposite trend. Irrespective of the sampling method, the dominance, evenness, shannon and simpson’s diversity indices were higher in the infested area for both foliage and ground-active arthropods. However, the Chao-1 estimator for family richness showed mixed results: In foliage samples, site 1 had a notably higher Chao-1 value compared to site 2. Conversely, ground samples revealed a higher Chao-1 value in site 1 (46) compared to Site 2 (37), whereas the overall Chao-1 estimates were higher for site 1 than site 2.
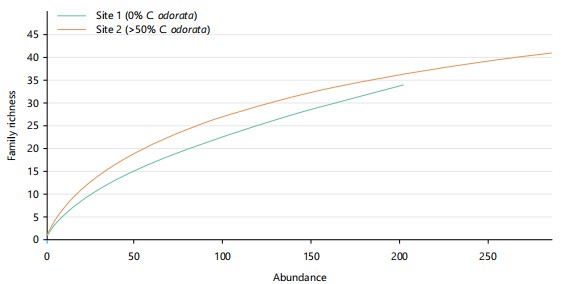
The rarefaction graph shows family richness across the two sites (0 and >50%). Each curve represents the cumulative family richness with increased sampling effort. Site 1, with 0% C. odorata invasion, shows lower family richness while site 2, with >50% C. odorata invasion, shows the lowest family richness. However, both curves are ascending, which indicates insufficient sampling depth or effort, hence, more sampling will lead to the discovery of new species as shown in Fig. 1.
| Table 1: | Arthropod families collected using pitfall traps and beating trays in sites infested without C. odorata (site 1) and that with infestation (site 2) | |||
| Family | Site 1 | Site 2 | Abundance | R.A (%) |
| Blaberidae | 0 | 1 | 1 | 0.2 |
| Bruchidae | 0 | 1 | 1 | 0.2 |
| Calliphoridae | 0 | 3 | 3 | 0.61 |
| Carabidae | 8 | 16 | 24 | 4.9 |
| Cerambycidae | 1 | 2 | 3 | 0.61 |
| Ceratopogonidae | 1 | 0 | 1 | 0.2 |
| Chrysomelidae | 1 | 1 | 2 | 0.41 |
| Coccinellidae | 0 | 2 | 2 | 0.41 |
| Crabronidae | 4 | 5 | 9 | 1.8 |
| Curculionidae | 3 | 10 | 13 | 2.7 |
| Ctenidae | 0 | 2 | 2 | 0.41 |
| Cydnidae | 1 | 4 | 5 | 1.02 |
| Cyrtaucheniidae | 1 | 1 | 2 | 0.41 |
| Dinidoridae | 1 | 0 | 1 | 0.2 |
| Ectobiidae | 2 | 0 | 2 | 0.41 |
| Elateridae | 1 | 0 | 1 | 0.2 |
| Endomychidae | 0 | 2 | 2 | 0.41 |
| Evaniidae | 0 | 36 | 36 | 7.4 |
| Forficulidae | 0 | 2 | 2 | 0.41 |
| Formicidae | 102 | 81 | 183 | 37.5 |
| Gryllidae | 8 | 0 | 8 | 1.64 |
| Glossinidae | 0 | 5 | 5 | 1.02 |
| Gnaphosidae | 0 | 15 | 15 | 0.31 |
| Hodotermopsidae | 0 | 1 | 1 | 0.2 |
| Lampyridae | 1 | 4 | 5 | 1.02 |
| Linyphiidae | 1 | 2 | 3 | 0.61 |
| Lycosidae | 1 | 0 | 1 | 0.2 |
| Meinertellidae | 2 | 3 | 5 | 1.02 |
| Melolonthinae | 1 | 0 | 1 | 0.2 |
| Membracidae | 0 | 1 | 1 | 0.2 |
| Muscidae | 16 | 7 | 23 | 4.7 |
| Nemonychidae | 1 | 0 | 1 | 0.2 |
| Nitidulidae | 1 | 7 | 8 | 1.64 |
| Oniscidae | 0 | 1 | 1 | 0.2 |
| Pentatomidae | 1 | 1 | 2 | 0.41 |
| Pholcidae | 1 | 0 | 1 | 0.2 |
| Plataspidae | 0 | 2 | 2 | 0.41 |
| Pyrgomorphidae | 0 | 1 | 1 | 0.2 |
| Pyrrhocoridae | 1 | 0 | 1 | 0.2 |
| Reduviidae | 1 | 3 | 4 | 0.82 |
| Salticidae | 1 | 8 | 9 | 1.84 |
| Scarabaeidae | 14 | 10 | 24 | 4.92 |
| Segestriidae | 0 | 1 | 1 | 0.2 |
| Spirostreptidae | 4 | 4 | 8 | 1.64 |
| Staphylinidae | 2 | 0 | 2 | 0.41 |
| Stratiomyidae | 0 | 1 | 1 | 0.2 |
| Tenebrionidae | 2 | 1 | 3 | 0.61 |
| Tetrigidae | 0 | 1 | 1 | 0.2 |
| Tettigoniidae | 1 | 2 | 3 | 0.61 |
| Theridiosomatidae | 10 | 1 | 11 | 2.25 |
| Thyreocoridae | 0 | 4 | 4 | 0.82 |
| Tingidae | 0 | 3 | 3 | 0.61 |
| Titanoecidae | 1 | 0 | 1 | 0.2 |
| Unidentified | 5 | 28 | 33 | 6.76 |
| R.A: Relative abundance, Site 1: Uninfested site (0% C. odorata) and Site 2: Infested site (>50% C. odorata) | ||||
|
| Table 2: | Family level diversity indices of arthropods sampled in site 1 and 2 | |||
| Ground arthropods | Foliage arthropods | Overall | ||||
| Indices | Site 1 | Site 2 | Site 1 | Site 2 | Site 1 | Site 2 |
| Family richness | 15 | 17 | 24 | 32 | 34 | 42 |
| Individuals | 81 | 61 | 121 | 225 | 202 | 286 |
| Dominance_D | 0.5428 | 0.326 | 0.1662 | 0.0979 | 0.2741 | 0.1183 |
| Simpson_1-D | 0.4572 | 0.674 | 0.8338 | 0.9021 | 0.7259 | 0.8817 |
| Shannon_H | 1.202 | 1.864 | 2.374 | 2.791 | 2.153 | 2.804 |
| Evenness_e^H/S | 0.2218 | 0.3794 | 0.4473 | 0.5092 | 0.2533 | 0.393 |
| Chao-1 | 48 | 21 | 46 | 37 | 72 | 52.11 |
DISCUSSION
The effect of C. odorata invasion on arthropod communities has important consequences for nature conservation and assessing potential ecosystem impacts. This study examined this effect by examining arthropod abundance and diversity using various diversity indices. At a family level, the current study compared the abundance and diversity of arthropods between an area with no C. odorata infestation (0%) and one with over >50% C. odorata coverage. Contrary to the expectations that arthropod abundance and diversity would be lower in the >50% C. odorata site, the results revealed higher dominance, diversity, abundance and evenness in the infested area. These findings contrast with those of van Hengstum et al.27, who reported that arthropod communities in invaded habitats tend to be smaller and less diverse and Mgobozi et al.24, observed lower diversity and species richness of spiders in C. odorata-infested sites compared to uninfested ones. The C. odorata-infested area showed an increased abundance of certain spider families, including Salticidae and Gnaphosidae, in comparison to the uninfested site. This finding contradicts Mgobozi et al.24, who observed that C. odorata infestation led to a decrease in the number of spiders and altered spider community patterns, species richness and diversity. However, they also found that the arthropod community recovered after clearing the invasion.
The 51% similarity, as shown by the Jaccard index, indicates a moderate overlap in arthropod family composition between the two sites, with approximately half of the families shared. This suggests that while C. odorata infestation modifies the structure of the community, it does not completely transform family composition, indicating that certain families may have adaptive traits that allow them to persist in infested environments. Generally, plant invasions are thought to reduce native plant richness. Since arthropods often depend on these plants for food and shelter and as sites to rear their young, the invasions may have positively impacted their populations36. The higher arthropod diversity observed in the C. odorata-infested area could be due to the additional shelter provided by C. odorata, which might attract arthropods that feed on surrounding native plants. The hypothesis that this increased shelter availability could lead to a larger and more diverse arthropod community in the >50% C. odorata site remains unproven but offers a plausible explanation.
The focus on arthropod communities at the family level provided a broad view of the composition; studies conducted at species or genus levels may reveal more intricate differences in arthropod composition between infested and uninfested sites37,38. For example, it is plausible that there would be high taxonomic variations from the samples collected, which in turn may or may not outweigh the functional relevance of the resident species. Furthermore, the current study also recognizes that pitfall and beating tray sampling may only capture part of the arthropod community, as shown by the rarefaction curves and some variations may be missed in the samples. For example, in dense C. odorata infestations, fast-flying insects could quickly leave the beating tray and some small arthropods may escape detection, especially in thick vegetation.
CONCLUSION
This study highlights the impact of C. odorata invasion on arthropod communities by assessing their community composition across infested and uninfested sites. Contrary to expectations of reduced arthropod diversity and abundance in infested areas, the site with over >50% C. odorata coverage showed higher arthropod family richness, diversity and evenness compared to the uninfested site. This finding suggests that C. odorata invasion may enhance certain aspects of arthropod community structure, potentially due to the additional shelter and complex habitat provided by the invasive plant. The study has shown an overlap in family composition between the two sites, indicating that while C. odorata modifies community structure, it does not entirely displace native arthropod families. Further research on the invasion duration of C. odorata and species-level responses would be valuable to deepen the understanding of the ecological effects of this invasive plant. Nonetheless, these findings contribute important insights to ongoing discussions on faunal community compositional responses to C. odorata invasion.
SIGNIFICANCE STATEMENT
Chromolaena odorata displaces native plants and modifies plant community structure. However, there is limited knowledge of its impact on arthropods associated with native flora. This study assessed the effect of C. odorata invasion on arthropod community assemblage. Samples were collected using pitfall traps and beating tray (foliage beating) methods. Current study results revealed that the presence of C. odorata increased family diversity, richness and evenness, probably due to the shaded habitat it provides when compared with sites without C. odorata. Further research on species-level responses would be valuable to deepen the understanding of the ecological effects of this invasive plant.
ACKNOWLEDGMENTS
We thank Efosa Edugie, Oshooluwa Jayeoba and Samuel Akinola for their invaluable assistance during the fieldwork. The current study also extend the thanks to Ogua community for providing the research site and to the University of Benin and Federal University Wukari for their immense support throughout the research.
REFERENCES
- Timbilla, J.A., C. Zachariades and H. Braimah, 2003. Biological Control and Management of the Alien Invasive Shrub Chromolaena odorata in Africa. In: Biological Control in IPM Systems in Africa, Neuenschwander, P., C. Borgemeister and J. Langewald (Eds.), CABI International, Wallingford, United Kingdom, ISBN: 978-1-84593-338-8, pp: 145-160.
- Zahara, M., 2019. Description of Chromolaena odorata L. R.M King and H. Robinson as medicinal plant: A review. IOP Conf. Ser.: Mater. Sci. Eng., 506.
- Muniappan, R., G.V.P. Reddy and P.Y. Lai, 2005. Distribution and Biological Control of Chromolaena odorata. In: Invasive Plants: Ecological and Agricultural Aspects, Inderjit (Ed.), Birkhäuser Basel, Basel, Switzerland, ISBN: 978-3-7643-7380-1, pp: 223-233.
- Li, Y.P., Y.L. Feng, W.T. Li, K. Tomlinson, Z.Y. Liao, Y.L. Zheng and J.L. Zhang, 2022. Leaf trait association in relation to herbivore defense, drought resistance, and economics in a tropical invasive plant. Am. J. Bot., 109: 910-921.
- Olawale, F., K. Olofinsan and O. Iwaloye, 2022. Biological activities of Chromolaena odorata: A mechanistic review. South Afr. J. Bot., 144: 44-57.
- Catarino, L., B. Indjai, M.C. Duarte and F. Monteiro, 2019. Chromolaena odorata invasion in Guinea-Bissau (West Africa): First records and trends of expansion. BioInvasions Rec., 8: 190-198.
- Robertson, M.P., M.H. Villet, D.H.K. Fairbanks, L. Henderson and S.I. Higgins et al., 2003. A proposed prioritization system for the management of invasive alien plants in South Africa: Research in action. S. Afr. J. Sci., 99: 37-43.
- Kriticos, D.J., T. Yonow and R.E. McFadyen, 2005. The potential distribution of Chromolaena odorata (Siam weed) in relation to climate. Weed Res., 45: 246-254.
- Ivens, G.W., 1974. The problem of Eupatorium odoratum L. in Nigeria. PANS Pest Art. News Summaries, 20: 76-82.
- Lucas, E.O., 1989. Siam weed (Chromolaena odorata) and crop production in Nigeria. Outlook Agric., 18: 133-138.
- Uyi, O.O., F. Ekhator, C.E. Ikuenobe, T.I. Borokini and E.I. Aigbokhan at el., 2014. Chromolaena odorata invasion in Nigeria: A case for coordinated biological control. Manage. Biol. Invasions, 5: 377-393.
- Uyi, O.O., 2020. Light environments affect herbivory patterns but not reproductive performance of a multivoltine specialist moth, Pareuchaetes pseudoinsulata. Sci. Rep., 10.
- Zachariades, C., M. Day, R. Muniappan and G.V.P. Reddy, 2009. Chromolaena odorata (L.) King and Robinson (Asteraceae). In: Biological Control of Tropical Weeds Using Arthropods, Muniappan, R., G.V.P. Reddy and A. Raman (Eds.), Cambridge University Press, Cambridge, England, ISBN: 9780511576348, pp: 130-162.
- Ikuenobe, C.E. and G.O. Anoliefo, 2003. Influence of Chromolaena odorata and Mucuna pruriens fallow duration on weed infestation. Weed Res., 43: 199-207.
- Adebayo, R.A. and U.O. Uyi, 2010. Biological control of invasive weed species: Nigerian experience. Int. J. Agric. Res., 5: 1100-1106.
- Uyi, O.O., C. Zachariades and M.P. Hill, 2014. The life history traits of the arctiine moth Pareuchaetes insulata, a biological control agent of Chromolaena odorata in South Africa. Afr. Entomol., 22: 611-624.
- Aigbedion-Atalor, P.O., M. Adom, M.D. Day, O. Uyi and I.N. Egbon et al., 2019. Eight decades of invasion by Chromolaena odorata (Asteraceae) and its biological control in West Africa: The story so far. Biocontrol Sci. Technol., 29: 1215-1233.
- Wei, H., J. Xu, G. Quan, J. Zhang and Z. Qin, 2017. Invasion effects of Chromolaena odorata on soil carbon and nitrogen fractions in a tropical savanna. Ecosphere, 8.
- Agboola, O.O. and J.I. Muoghalu, 2015. Changes in species diversity, composition, growth and reproductive parameters of native vegetation invaded by Chromolaena odorata and Tithonia diversifolia in Osun State, Southwest Nigeria. FUTA J. Res. Sci., 11: 217-230.
- Cripps, M.G., M. Schwarzländer, J.L. McKenney, H.L. Hinz and W.J. Price, 2006. Biogeographical comparison of the arthropod herbivore communities associated with Lepidium draba in its native, expanded and introduced ranges. J. Biogeogr., 33: 2107-2119.
- Effah, E., D.P. Barrett, P.G. Peterson, M.A. Potter, J.K. Holopainen and A.C. McCormick, 2020. Effects of two invasive weeds on arthropod community structure on the Central Plateau of New Zealand. Plants, 9.
- Emery, S.M. and P.J. Doran, 2013. Presence and management of the invasive plant Gypsophila paniculata (baby’s breath) on sand dunes alters arthropod abundance and community structure. Biol. Conserv., 161: 174-181.
- Costello, S.L., P.D. Pratt, M.B. Rayamajhi and T.D. Center, 2003. Arthropods associated with above-ground portions of the invasive tree, Melaleuca quinquenervia, in South Florida, USA. Fla. Entomol., 86: 300-322.
- Mgobozi, M.P., M.J. Somers and A.S. Dippenaar-Schoeman, 2008. Spider responses to alien plant invasion: The effect of short- and long-term Chromolaena odorata invasion and management. J. Appl. Ecol., 45: 1189-1197.
- Rizali, A., M.S. Hadi, Pudjianto and D. Buchori, 2019. A new trophic interaction between invasive weed, its biological control agent, and local insects: A case study of Chromolaena odorata. Biodiversitas J. Biol. Diversity, 20: 1006-1011.
- Buchori, D., A. Rizali, L. Lukvitasari and H. Triwidodo, 2020. Insect communities associated with siam weed: Evaluation after three decades of Cecidochares connexa release as biocontrol agent. Diversity, 12.
- van Hengstum, T., D.A.P. Hooftman, J.G.B. Oostermeijer and P.H. van Tienderen, 2014. Impact of plant invasions on local arthropod communities: A meta-analysis. J. Ecol., 102: 4-11.
- Roper-Edwards, I.R. and A.H. Hurlbert, 2024. Arthropod community composition in urban landscapes is shaped by both environmental filtering and dispersal limitation. PLoS ONE, 19.
- Litt, A.R., E.E. Cord, T.E. Fulbright and G.L. Schuster, 2014. Effects of invasive plants on arthropods. Conserv. Biol., 28: 1532-1549.
- Lauchande, V.L., S.P. Mntambo, Z.T. Hlongwane and T.C. Munyai, 2024. Comparing the effectiveness of pitfall traps and active sampling methods for ants and spiders in a Chromolaena odorata invaded site. Bothalia-Afr. Biodivers. Conserv., 54.
- Oliver, I. and A.J. Beattie, 1996. Designing a cost-effective invertebrate survey: A test of methods for rapid assessment of biodiversity. Ecol. Appl., 6: 594-607.
- Borror, D.J., C.A. Triplehorn and N.F. Johnson, 1989. An Introduction to the Study of Insects. 6th Edn., Saunders College Pub., Philadelphia, ISBN: 9780030253973, Pages: 875.
- Hohbein, R.R. and C.J. Conway, 2018. Pitfall traps: A review of methods for estimating arthropod abundance. Wildl. Soc. Bull., 42: 597-606.
- Hammer, Ø. and D.A.T. Harper, 2024. Paleontological Data Analysis. 2nd Edn., John Wiley & Sons, Hoboken, New Jersey, ISBN: 978-1-119-93395-3, Pages: 400.
- Konopiński, M.K., 2020. Shannon diversity index: A call to replace the original Shannon’s formula with unbiased estimator in the population genetics studies. PeerJ, 8.
- Spafford, R.D., C.J. Lortie and B.J. Butterfield, 2013. A systematic review of arthropod community diversity in association with invasive plants. NeoBiota, 16: 81-102.
- Rosser, N. and P. Eggleton, 2012. Can higher taxa be used as a surrogate for species-level data in biodiversity surveys of litter/soil insects? J. Insect Conserv., 16: 87-92.
- Rosser, N., 2017. Shortcuts in biodiversity research: What determines the performance of higher taxa as surrogates for species? Ecol. E, 7: 2595-2603.
How to Cite this paper?
APA-7 Style
Otabor,
J.I., Udemadu,
M.O., Osawe,
N., Isikhuemen,
C.O., Okafor,
M.N., Uyi,
O. (2025). Invasive Chromolaena odorata Infestation Supports Increased Arthropod Diversity in Southern Nigeria. Asian Journal of Biological Sciences, 18(1), 237-245. https://doi.org/10.3923/ajbs.2025.237.245
ACS Style
Otabor,
J.I.; Udemadu,
M.O.; Osawe,
N.; Isikhuemen,
C.O.; Okafor,
M.N.; Uyi,
O. Invasive Chromolaena odorata Infestation Supports Increased Arthropod Diversity in Southern Nigeria. Asian J. Biol. Sci 2025, 18, 237-245. https://doi.org/10.3923/ajbs.2025.237.245
AMA Style
Otabor
JI, Udemadu
MO, Osawe
N, Isikhuemen
CO, Okafor
MN, Uyi
O. Invasive Chromolaena odorata Infestation Supports Increased Arthropod Diversity in Southern Nigeria. Asian Journal of Biological Sciences. 2025; 18(1): 237-245. https://doi.org/10.3923/ajbs.2025.237.245
Chicago/Turabian Style
Otabor, Jane, I., Mildred O. Udemadu, Nosa Osawe, Cornell O. Isikhuemen, May N. Okafor, and Osariyekemwen Uyi.
2025. "Invasive Chromolaena odorata Infestation Supports Increased Arthropod Diversity in Southern Nigeria" Asian Journal of Biological Sciences 18, no. 1: 237-245. https://doi.org/10.3923/ajbs.2025.237.245

This work is licensed under a Creative Commons Attribution 4.0 International License.