Differential Biotic and Abiotic Induced Stress Levels Confer Varying Morphological Responses in Okra (Abelmoschus esculentus L. Moench) Cultivars
-
Ayisha Marfo Amadu
Department of Plant and Environmental Biology, School of Biological Sciences, University of Ghana-Legon, P.O. box LG 55 Legon, Ghana
Andrew Sarkodie AppiahBiotechnology and Nuclear Agriculture Institute, Ghana Atomic Energy Commission, Kwabenya, Ghana
Kwadwo Owusu Ayeh
Department of Plant and Environmental Biology, School of Biological Sciences, University of Ghana-Legon, P.O. box LG 55 Legon, Ghana
| Received 15 Mar, 2023 |
Accepted 15 Jun, 2023 |
Published 30 Sep, 2023 |
Background and Objective: Extreme and prolonged water deficit together with the evolving nature of plant viruses has a negative impact on plant growth. This study was thus conducted to evaluate the morphological responses of okra cultivars subjected to drought and virus stress treatments. Materials and Methods: Three watering treatment blocks were set up in a split block design and plants under each block were arranged further in a Randomized Complete Block Design (RCBD). Means of plant growth measurements and yield parameters were compared using One-way Analysis of Variance (ANOVA) statistics using Minitab (version 17.0) statistical software and means of treatments were compared by Tukey mean comparison at a 5% probability level. Results: The lowest reduced plant height of 19.60±0.84 cm corresponded to Indiana plants grown under a combination of severe drought and virus treatment. The least number of leaves were reported in Indiana plants grown under a combination of severe drought and virus treatment (1.66±0.57). The highest and least stem diameter were recorded in Essoumtem control plants (9.10±0.17 mm) and Indiana plants grown under a combination of severe drought and virus treatment (2.96±0.15 mm), respectively (p = 0.00, F = 74.53). Conclusion: Results concluded that virus infection and drought stressed plants may result in impaired growth in okra cultivars used in this study.
| Copyright © 2023 Amadu et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Okra is one of the crops that have the potential of improving food security and malnutrition and alleviating poverty in sub-tropical and the tropical regions of the world. In addition, Okra constitutes a greater percentage of fresh vegetables, exported from India to Europe and the Middle East which serves as one of the global market place for the consumption of Okra1. Although the crop is high yielding under improved production practices, several stresses negatively affect the growth, yield and biological activities of the plant worldwide2,3. Specifically, okra is affected by several plant viruses such as the yellow vein mosaic virus and enation leaf curl virus which leads to great economic losses4.
Plant stress is a state where a plant is grown under unfavourable growth conditions affecting its entire morphogenetic growth and developmental processes. This situation puts the plant under increasing demand for water and nutritional needs which if not attained over a prolonged period leads to senescence of the plant. They adapt to these stress conditions and minimize their effect by adjusting their physiology, morphology and sometimes metabolism5. Plant stresses are generally considered either biotic (those caused by biological agents) or abiotic (those resulting from the physical environment). They have negative effects on cell growth, cause separation of membrane proteins, decrease chlorophyll content in leaves and may sometimes result in loss of germination capability6. Biotic stress in plants is caused by living organisms, specifically viruses, fungi, bacteria, insects, nematodes, weeds and arachnids. In response to these biotic stress factors, plants have evolved sophisticated mechanisms to fight biotic stress including the production of several biotic stress resistance genes7. In addition to biotic stress, plants may be exposed to abiotic stress as well as a combination of both8. When there is a concurrent infection of an abiotic and biotic stresses on a plant, a situation is created where symptoms produced may become aggravated or lessen the effects of one of the double infection. Rivero et al.9 revealed that, when plants are exposed to abiotic stress factor, there is a net worsening effect of an introduced abiotic stress such as water stress factor which could reduce the plant’s ability to defend itself against the earlier introduced pathogen. Gupta et al.10, however mentioned that the outcome of double stress interactions varies depending on timing, the severity of the stresses (drought and pathogens), the order of occurrence of each stressor and the intensity and duration of exposure. Viruses as biotic stress factors are genetic elements that cannot replicate independently outside a living host cell.
Simultaneous occurrence of drought and pathogen stresses is considered one of the most important combinations affecting crops worldwide11,12. This was because, plants growing under stressful conditions such as drought may be weak in protecting themselves and pathogens can easily attack such plants and cause further damage to them. Suffice it to say, when there is a coexistence of both biotic and abiotic stresses on a plant, the net result may be either mild or severe symptoms on plants which could also depend on the pathogen or the imposed stress factor on the plant13. Moreover, a plant’s response to water deficit may meddle with plant-virus infection in view of the fact that viral infection can safeguard the host plant’s susceptibility by delaying permanent wilting in a water-deficit environment14. With continuous changes in climatic conditions, the occurrence of combined biotic and abiotic stresses on plants is likely to increase albeit very little understanding of the morphological development that occurs in plants experiencing a combination of biotic and abiotic stresses is known. It is therefore necessary to evaluate the effects of simultaneous biotic and abiotic stresses on plant growth and development. There is therefore the need to understand the morphological changes associated with these biotic and abiotic stresses on okra cultivars to deepen the understanding of how these stresses affect plant growth.
MATERIALS AND METHODS
Study area: The experiment was carried out in the screen house of the Department of Plant and Environmental Biology, University of Ghana at a temperature of 35°C and 40% relative humidity. The duration of the experiment was from September, 2020 to June, 2021.
Plant material: Three commercially grown cultivars of okra (Abelmoschus esculentus) namely, Indiana, Essoumtem and Rafiki were obtained from Agriseed Ltd., Technisem Ghana and used for this study (Fig. 1a-c). Infected okra leaves used for mechanical sap inoculation were collected from an okra field at Ghana Atomic Energy Commission (GAEC).

|
Soil: Humus soil, used for the study was obtained from the Teaching Garden of the Department of Plant and Environmental Biology, University of Ghana and sterilized using an autoclave at a pressure of 15 psi and temperature of 121°C for 30 min. Two kilograms of soil was each weighed and placed in polythene bags of dimension 20×30 cm. A total of 117 bags were used for the study.
Planting: Perforated polythene bags were filled with sterilized soil and completely saturated with 300 mL of water a day prior to planting seeds. Three seeds of each cultivar were planted in separate polythene bags and rehydrated with 200 mL of water, followed by 100 mL of water every other day.
Buffer preparation: Buffer used for mechanical inoculation was prepared following the Q-bank plant viruses and viroids protocol. Phosphate buffer (10X) was prepared by dissolving 0.272 g of KH2PO4 and 1.42 g of Na2HPO4×2H2O in 100 mL of demineralized water pH 7.4 using NaOH. Ten grams of polyvinylpyrrolidone (PVP, MW 10,000) was weighed and added to the phosphate buffer and 400 mL of demineralized water was added to a final volume of 500 mL of inoculation buffer.
Preparation of inoculum and virus inoculation: Plants were thinned out after 14 days post-planting, leaving one in each bag. Older leaves of plants were removed, leaving two leaves on each plant. Plants were covered with black polythene overnight and removed just before the mechanical inoculation was carried out.
Virus-infected okra leaves (source of inoculum) were obtained from planting fields of GAEC, separately weighed and ground using sterilized mortars and pestles. One gram of ground infected okra leaves was added to 10 mL of inoculation buffer with celite and mixed thoroughly. Fifteen plants of each cultivar were inoculated by rubbing the mixture on uninfected leaves. Inoculated leaves were immediately washed off with distilled water.
Experimental design: Three watering treatment blocks (normal watering, moderate watering and severe drought) were set up in a split block design and plants under each block were arranged in Randomized Complete Block Design (RCBD). Each block was further divided into three subplots with treatments that included virus stress, water stress, a combination of both virus and water stresses and control plants.
Experimental set-up: Virus-inoculated and virus-free seedlings were then placed under the treatment blocks as follows: Each sub-treatment had 5 replicates of each cultivar except control plants which were in replicates of three giving a total of 9 control plants, 15 water-stressed plants and 15 water plus virus stressed plants per treatment blocks (Normal, Moderate and Severe). The different watering regimes were induced after the introduction of virus stress so all stresses could be monitored simultaneously.
Watering regimes: Three sets of water regimes were used throughout the experimental setup. These included normal watering, moderate watering and severe drought treatments where plants were rehydrated every other day, every 3 and 5 days, respectively. Control plants were however rehydrated daily. All plants were rehydrated with 150 mL of water.
Data collection
Virus symptomatology on leaves: After 7 days of inoculation, all five replicates per cultivar in all virus-inoculated treatment blocks were monitored for 10 consecutive weeks for virus symptoms using a scale of 0-5:
| 0 | = | Healthy, asymptomatic plant | |
| 1 | = | Mild mosaic patches | |
| 2 | = | Moderate mosaic patches, yellowing of leaf | |
| 3 | = | Yellowing of leaf vein, mosaic, yellowing of leaf, mild stunted growth | |
| 4 | = | moderate chlorosis, yellowing of leaf vein, mosaic with leaf abscission, stunted growth | |
| 5 | = | Severe chlorosis, mosaic, leaf abscission, yellowing of leaf vein and stunted growth |
Measurement of growth and development parameters.
Plant height: Plant height was measured from the soil surface to the terminal bud at the apex of the shoot using a meter rule for 10 consecutive weeks.
Number of leaves: Mean number of leaves per plant was counted from 5 replicates used in each treatment.
Internode length: The length of the third internode from the base of each plant was measured using a meter rule and the diameter of each stem was measured using a digital calliper (Neiko 01407a electronic digital calliper 150 mm).
Leaf area: The leaf area of the second and third leaves from the apex of each plant was used for measuring leaf area using the formula (Eq. 1):
Leaf length×Leaf width×Leaf area coefficient |
(1) |
where, the leaf area coefficient of okra used in this work was 0.62 according to Musa and Usman15. An average of the two leaves were calculated per plant.
Statistical analysis: Means of plant growth measurements were compared using One-way Analysis of Variance (ANOVA) statistical software (Minitab version 17.0). Means of treatments were compared by Tukey mean comparison method at a 5% probability level and values were expressed as Mean±SE (standard error). Correlational analysis of morphological growth was carried out using Stata (version 15) statistical analysis software to show the relationship among the traits studied.
RESULTS
Morphological/plant growth measurements
Plant height: Plant heights in the Rafiki cultivar, compared at a juvenal stage of 28 DAI, showed that plants under both control and normal watering regimes were significantly different from plants infected with the virus (p = 0.00, F= 10.05). The highest mean plant height corresponded to 38.52±2.51 cm in the control water treatment. This was not significantly different from normal and moderate water treatment but significantly different from the rest of the treatments in this study (Table 1). Rafiki plants there were no significant differences in plant height in control, normal and moderate watering regimes (Fig. 2a-c). However, plant height in severe drought treatment without virus inoculation (Fig. 2d) was significantly different from plant height in control, normal and moderate watering regimes. Interestingly plant height in moderate water plus virus inoculation treatments (2E) was significantly not different from control, normal and moderate watering regimes. The two least plant heights were found in moderate watering with a virus (2F) and severe watering plus virus (2G). However, they were not significantly different from severe drought treatment.
When Indiana cultivars under the various biotic and abiotic treatments were compared at 28 DAI, control plants had the highest plant height of 35.06±5.63 cm and varied significantly among all other treatments except plants under normal and moderate watering (P = 0.00, F = 8.06). The least plant height of 18.30±0.56 cm was recorded in Indiana plants under a combination of severe drought and virus stresses (pictures not shown). This was however not significantly different from plants subjected to moderate watering, combined effect of moderate watering and virus infection, combination of normal watering and virus infection and severe drought treatments (Table 1).
At 28 DAI, Essoumtem control plants had the highest plant height (34.44±1.02 cm) (Table 1) and it was significantly different from plants subjected to moderate watering, a combination of moderate watering and virus stress, severe drought and a combination of severe drought and virus stress (P = 0.00, F = 7.53). The least plant height of 26.14±2.18 cm was seen in plants under a combination of severe drought and virus stress and it varied significantly from control plants and those under a combination of normal watering and virus stress.

|
| Table 1: | Plant height of three cultivars under three watering regimes and a combination of watering regimes and virus stress at juvenile age (28 DAI) | |||
Mean plant height at 28 DAI±SE (cm) |
|||||||
| Cultivar | Control | Normal | Normal+virus | Moderate | Moderate+virus | Severe | Severe+virus |
| Rafiki | 38.52±2.51a | 36.16±2.46ab | 26.42±4.34def | 31.04±3.74abcde | 20.20±0.96f | 24.94±7.43ef | 25.83±3.88cdef |
| Indiana | 35.06±5.63abc | 33.86±1.84ab | 23.48±1.20ef | 26.66±6.61cdef | 23.50±1.13def | 23.30±3.82ef | 18.30±0.56f |
| Essoumtem | 34.44±1.02abcd | 30.58±2.59abcde | 31.30±3.65abcde | 27.30±2.20cdef | 28.60±0.57bcdef | 28.16±2.10bcdef | 26.14±2.18def |
| Means that do not share a letter are significantly different at a 95% confidence Interval | |||||||
| Table 2: | Plant height of three cultivars under three watering regimes and a combination of watering regimes and virus stress at maturity (70 DAI) | |||
Mean plant height at 70 DAI±SE (cm) |
|||||||
| Cultivar | Control | Normal | Normal+virus | Moderate | Moderate+virus | Severe | Severe+virus |
| Rafiki | 89.60±5.30a | 81.96±2.96ab | 66.32±10.25cd | 54.04±8.41de | 44.33±4.31ef | 47.78±10.07ef | 40.13±7.03efgh |
| Indiana | 51.10±3.83ef | 39.94±3.78fg | 28.50±5.15ghi | 28.42±6.78ghi | 23.10±3.82ghi | 25.65±4.56hi | 19.60±0.84i |
| Essoumtem | 76.22±3.40bc | 65.48±2.04cd | 64.22±6.75cd | 53.86±2.42de | 50.30±3.38ef | 44.78±1.64ef | 43.58±1.24ef |
| Means that do not share a letter are significantly different at a 95% confidence interval | |||||||
At maturity (70 DAI), it was observed in Rafiki cultivars that, the higher the stress treatment, the lower the plant height. Control plants and plants under normal watering recorded the highest heights (89.60±5.30 and 81.96±2.96 cm, respectively) and were significantly different from plants under all other treatments (P = 0.00, F= 27.48). Plants under severe drought, a combination of moderate watering and virus stress and under a combination of severe drought and virus stresses recorded the least plant heights (47.78±10.07, 44.33±4.31 and 40.13±7.03 cm), respectively but were not significantly different from each other even though there were differences in their heights (Table 2).
At 70 DAI, the highest plant height was seen in Indiana control plants (51.10±3.83 cm) and was significantly different from plants under all other treatments (P = 0.00, F = 20.67). The least plant height of 19.60 cm±0.84 was observed in Indiana plants under a combination of severe drought and virus stresses but was however not significantly different from plants under all treatment levels except control plants and plants under normal watering (Table 2). When the plant height of the Essoumtem cultivar was compared among the different stress treatments at maturity (70 DAI), it was observed that generally, plant height decreased with increasing stress levels. The control plant had the highest height (76.22 cm±3.40) and it varied significantly from plants under all the other treatments (P= 0.00, F = 61.06). Plants under normal watering and a combination of normal watering and virus infection had the next highest plant heights and were significantly different from plants under all other stress treatments (Table 2).
Plants subjected to a combination of severe drought and virus infection recorded the least height (43.58±1.24 cm) and it varied significantly from plants under all other treatments except those under severe drought and a combination of moderate watering and virus infection. Rafiki cultivar under normal watering recorded the highest plant height of 81.96±2.96 cm. However, under a combination of normal watering and virus treatment, plant height was impacted and reduced to 66.32±10.25 cm and this was significantly different from Rafiki plants under normal watering and significantly different from the effect of combined normal water treatment and virus stresses in Essoumtem and Indiana cultivars (P = 0.00, F = 56.80). Indiana cultivar under a combination of both stresses recorded the least plant height (28.50±5.15 cm) and was significantly different from all plants under both stresses.
Generally, when all three cultivars were compared across all stress treatments, the least percentage height reduction was recorded in Rafiki under normal watering (6.27±3.38%). It varied significantly from plants under all the other treatments except Indiana and Essoumtem under normal watering and Rafiki and Essoumtem under a combination of normal watering and virus infection (P = 0.00, F = 15.56). The highest percentage of height reduction was seen in plants under a combination of severe drought and virus infection. Indiana and Rafiki cultivars under a combination of severe drought and virus infection had the highest percentage height reduction of 59.05±0.70 and 54.10±8.04%, respectively and were significantly different from all cultivars under normal watering, Essoumtem under moderate watering, Rafiki and Essoumtem under a combination of normal watering and virus infection and Essoumtem under a combination of moderate watering and virus infection (Table 3).
When cultivars were compared within treatments, it was observed in both Rafiki and Essoumtem cultivars that plants under a combination of severe drought and virus infection had the highest percentage height reductions and were significantly different (P = 0.00, F= 16.68 and P = 0.00, F = 37.01, respectively) from plants under normal watering and a combination of normal watering and virus infection which had the least plant height reduction (Table 3). Indiana plants under a combination of severe drought and virus infection had the highest percentage height reduction (59.05±0.70%) and it varied significantly from plants under normal watering only (P = 0.00, F = 7.83).
Number of leaves: Comparing among all treatments within the Rafiki cultivar, virus-inoculated plants under severe drought treatment had the least number of leaves (2.00±0.00) and it was significantly different from plants subjected to all the other treatments (F = 55.58, P = 0.00). The highest number of leaves was recorded in control plants and it varied significantly from the number of leaves recorded in all other treatments. Plants under moderate watering, severe drought, a combination of moderate watering and virus and a combination of normal watering and virus treatments recorded a varying number of leaves, but they did not vary significantly.
The highest number of leaves (10.00±1.22) corresponded to control plants in Essoumtem and this varied significantly (P = 0.00, F= 30.62) from plants under all other treatments except those subjected to normal watering (8.40±1.14) and a combination of normal watering and virus infection (8.40±0.54). The least number of leaves was recorded in plants under a combination of severe drought and virus infection (3.40±1.14). It was however not significantly different from plants subjected to moderate watering, severe drought and a combination of moderate watering and virus infection (Table 4).
Within Indiana cultivar, the least number of leaves was recorded in plants under a combination of severe drought and virus infection (1.66±0.57) (Table 4). It was however not significantly different from plants under all the other treatments even though they recorded a varying number of leaves except for control plants (7.80±0.83) (P = 0.00, F = 8.56).
When all three cultivars were compared among all treatments, Indiana plants subjected to a combination of severe drought and virus infection had the least mean number of leaves (Table 4). Control plants in Essoumtem had the highest number of leaves (10.0±1.22) and it varied significantly among all cultivars under all treatments except Indiana and Rafki controls, Essoumtem under normal watering and a combination of normal watering and virus infection (P = 0.00, F = 23.01).
| Table 3: | Percentage of plant height reduction in three okra cultivars under watering regimes and a combination of watering regimes and virus treatment at 70 DAI | |||
Percentage plant height reduction (%)±SE |
|||
Cultivar |
|||
| Treatment | Rafiki |
Indiana |
Essoumtem |
| Normal | 6.27±3.38g |
18.05±7.75efg |
11.75±2.76fg |
| Moderate | 38.20±9.61abcd |
41.69±13.91abcd |
27.41±3.26cdef |
| Severe | 45.36±11.52abc |
47.37±9.37ab |
39.65±2.22abcd |
| Normal+Virus | 24.15±11.72defg |
41.53±10.56abcd |
13.45±9.10efg |
| Moderate+Virus | 49.30±4.93ab |
51.44±1.97ab |
32.21±4.55bcde |
| Severe+Virus | 54.10±8.04ab |
59.05±0.70a |
41.26±1.68abcd |
| Means that do not share a letter are significantly different at 95% confidence interval | |||
| Table 4: | Mean number of leaves of three okra cultivars under three watering regimes and a combination of watering regimes and virus infections at 70 DAI | |||
Mean number of leaves±SE |
|||
Cultivar |
|||
| Treatment | Rafiki |
Indiana |
Essoumtem |
| Control | 8.20±0.83a |
7.80±0.83ab |
10.00±1.22a |
| Normal | 5.40±0.54bc |
3.80±1.64cde |
8.40±1.14a |
| Moderate | 3.40±0.54cde |
3.60±2.61cde |
4.80±0.83cd |
| Severe | 3.40±0.54cde |
1.75±1.70e |
4.80±0.44cd |
| Normal+Virus | 4.20±0.44cde |
2.80±1.09de |
8.40±0.54a |
| Moderate+Virus | 3.75±0.50cde |
2.75±0.50cde |
5.50±1.29bc |
| Severe+Virus | 2.00±0.00de |
1.66±0.57e |
3.40±1.14cde |
| Means that do not share a letter are significantly different at a 95% confidence interval | |||
| Table 5: | Means that do not share a letter are significantly different at a 95% confidence interval | |||
Mean symptom severity score±SE |
||||
| Cultivar | Control |
Normal watering+virus |
Moderate watering+virus |
Severe drought+virus |
| Rafiki | 0.00±0.00e |
1.60±0.54d |
3.00±0.00abc |
3.00±0.00abc |
| Indiana | 0.00±0.00e |
3.40±0.54ab |
4.00±0.00a |
4.00±0.00a |
| Essoumtem | 0.00±0.00e |
0.60±0.89e |
2.33±0.57cd |
3.00±0.00bc |
| Means that do not share a letter are significantly different at a 95% confidence interval | ||||
Effect of the virus on leaf morphology: Generally, a comparison within all three cultivars showed that high mean symptom severity scores of 3.00±0.00, 4.00±0.00 and 3.00±0.00 were recorded in virus-infected Rafiki, Indiana and Essoumtem plants subjected to severe drought. When all three cultivars were compared, the highest symptom severity score was recorded in Indiana plants under severe and moderate watering stresses and did not vary significantly from virus-infected plants under normal watering (Table 5). However, it varied significantly from virus-infected Rafiki plants subjected to normal watering and Essoumtem plants under all the different watering regimes (P=0.00, F= 24.44). Virus-infected Essoumtem plants under normal watering had the least mean symptom severity score of 0.60±0.89 and were significantly different from all cultivars under all the other watering regimes except control plants (Table 5). All control plants showed no virus symptoms and varied significantly from their respective diseased plants.
No visible symptoms were observed in Rafiki control (healthy) plants (Fig. 3a) as compared to typical virus symptoms such as mosaic patches on Rafiki leaves (Fig. 3b and c), yellowing of leaf veins, yellowing of leaves (chlorosis) and enations (Fig. 3d) were observed in Rafiki cultivars after 21 DAI.
No symptoms were recorded in Essoumtem control plants (Fig. 4a) as compared to yellowing of leaf veins and mosaic symptoms were the dominant virus symptoms observed on leaves of virus inoculated plants in Essoumtem (Fig. 4b-d).
No virus symptoms were observed in Indiana control cultivar (Fig. 5a) as compared to the more intense yellowing and clearing of leaf veins (Fig. 5b-d).
Internode length: When internode length was compared within the Rafiki cultivar, control plants had the highest internode length (6.56±0.83 cm) and varied significantly from all other treatments except plants under normal watering, moderate watering and severe drought (P = 0.02, F= 5.00). The least internode length of 2.66±0.43 cm was seen in plants under a combination of normal watering and virus infection and this varied significantly between control plants and plants subjected to severe drought (Table 6). In Indiana cultivar, the shortest internode length (0.85±0.35 cm) was recorded in plants under a combination of severe drought and virus treatment and it was significantly different from control plants (4.94±1.63 cm) and plants subjected to normal watering (P = 0.00, F = 7.76) (Table 6).

|

|

|
Comparison among all cultivars and treatments revealed that the highest (6.78±0.09 cm) and least (0.85±0.35 cm) internode lengths were recorded in Essoumtem control plants and Indiana plants under a combination of severe drought and virus infection and they were significantly different from each other (P = 0.00, F = 7.51). Rafiki and Indiana control plants, all plants under normal watering, Rafiki and Essoumtem plants under moderate watering and Rafiki and Essoumtem plants under severe drought also had high internode lengths and did not vary significantly from Essoumtem control plants (Table 6).
| Table 6: | Mean internode length of three okra cultivars under three watering regimes and a combination of watering regimes and virus infections at 70 DAI | |||
Mean internode length±SE |
|||
Cultivar |
|||
| Treatment | Rafiki |
Indiana |
Essoumtem |
| Control | 6.56±0.83a |
4.94±1.63abcd |
6.78±0.09a |
| Normal | 5.56±0.59abc |
4.46±0.64abcd |
4.76±1.02abcd |
| Moderate | 5.04±1.18abcd |
3.72±1.25bcdef |
5.98±0.50ab |
| Severe | 5.94±3.10ab |
2.67±1.15cdef |
5.52±0.87abc |
| Normal+virus | 2.66±0.43def |
1.46±0.38f |
4.18±0.79abcdef |
| Moderate+virus | 2.63±1.55cdef |
2.80±0.28bcdef |
4.15±1.26abcdef |
| Severe+virus | 2.76±1.65bcdef |
0.85±0.35ef |
4.38±0.42abcde |
| Means that do not share a letter are significantly different at a 95% confidence interval | |||
| Table 7: | Mean stem diameter of three okra cultivars under three watering regimes and a combination of watering regimes and virus infection at maturity (70 DAI) | |||
Mean stem diameter (mm)±SE |
|||
Cultivar |
|||
| Watering Regime | Rafiki |
Indiana |
Essoumtem |
| Control | 8.34±0.39ab |
6.82±0.57cd |
9.10±0.17a |
| Normal | 6.64±0.26de |
5.18±0.21gh |
7.80±0.40b |
| Moderate | 5.08±0.53ghi |
4.18±0.40ijk |
5.50±0.56fg |
| Severe | 4.91±0.20ghij |
3.42±0.45kl |
4.94±0.41ghij |
| Normal+virus | 6.22±0.23def |
4.20±0.50ijk |
7.66±0.54bc |
| Moderate+virus | 4.96±0.40ghij |
3.60±0.20kl |
5.65±0.73efg |
| Severe+virus | 4.13±0.11hijkl |
2.96±0.15l |
4.00±0.38jkl |
| Means that do not share a letter are significantly different at a 95% confidence interval | |||
Essoumtem control plants recorded the highest internode length (6.78±0.09 cm) and varied significantly from plants under a combination of moderate watering and virus infection which had the least internode length (4.15±1.26 cm). Essoumtem plants subjected to moderate watering and severe drought also had high internode lengths and were significantly different from plants under all treatments except control plants (P = 0.00, F = 6.77) (Table 6).
Stem diameter: At 70 DAI, a comparison among all three cultivars across all treatments showed that Essoumtem control plants recorded the highest stem diameter (9.10±0.17 mm). It varied significantly from plants under all treatments (P = 0.00, F = 74.53) except Rafiki control plants. Indiana plants under a combination of severe drought and virus infection had the least stem diameter (2.96±0.15 mm) and varied significantly from all other plants except Rafiki and Essoumtem plants under a combination of severe drought and virus infection, Indiana plants under severe drought and a combination of moderate watering and virus infection (Table 7).
Within Rafiki treatments, the highest (8.34±0.39 mm) and least (4.13±0.11 mm) stem diameter values were recorded in control plants and plants subjected to a combination of severe drought and virus infection respectively and they were significantly different from each other (P = 0.00, F = 76.02). Control plants varied significantly from plants under all other treatments (Table 7). The stem diameter of plants under a combination of severe drought and virus infection varied significantly from all other plants except those subjected to severe drought and a combination of moderate watering and virus infection (Table 7).
Generally, there were significant differences in stem diameter among treatments in Indiana cultivar at 70 DAI (P = 0.00, F = 43.55). The largest stem diameter (6.82±0.57 mm) was recorded in control plants and it was significantly different from plants under all other treatments. Plants under a combination of severe drought and virus infection had the least stem diameter (2.96±0.15 mm) and were significantly different from control plants, plants subjected to a combination of normal watering and virus infection, moderate watering and normal watering. Similar results were found in the Essoumtem cultivar (Table 7).
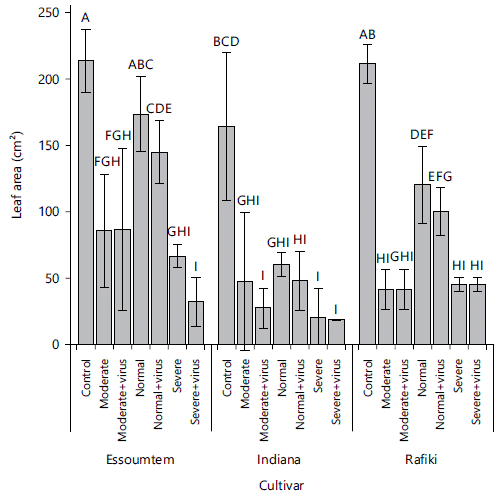
Leaf area: Generally, there were significant differences in leaf area among all three cultivars when compared to all stress treatments (P = 0.00, F = 40.00). The highest (213.92±18.88 cm2) and least (17.95±0.07 cm2) leaf area values were observed in Essoumtem control plants and Indiana plants under a combination of severe drought and virus infection, respectively and they varied significantly from each other (Fig. 6). Leaf area in Essoumtem was largest when compared among all three cultivars under all treatments. Indiana cultivar had the least leaf area under all treatments and varied from Rafiki and Essoumtem cultivars except when Essoumtem plants were subjected to moderate watering and Rafiki cultivar subjected to a combination of moderate and virus infection (Fig. 6).
Comparing within cultivar and among treatments, leaf area generally increased with decreasing stress level. Control plants in Rafiki had the highest leaf area (211.24±11.44 cm2) and it was significantly different from plants under all other treatments (P = 0.00, F = 106.44). A combination of severe drought and virus infection resulted in plants with the least leaf area (29.17±5.06 cm2) and they varied significantly from control plants, plants under normal watering and plants subjected to a combination of normal watering and virus infection (Fig. 6).
Root morphology and root to shoot ratio (r:s): The root to shoot ratio of three okra cultivars under three watering was shown in Table 8.
When all three cultivars were compared, Rafiki plants under severe drought had the highest (0.65±0.09) root to shoot ratio and whereas the least root to shoot ratio was recorded in Indiana control plants (0.15±0.02), varying significantly from each other (P = 0.00, F = 5.36).
|
| Table 8: | Root to shoot ratio of three okra cultivars under three watering regimes and a combination of watering regimes and virus infection at maturity (70 DAI) | |||
Mean root to shoot ratio±SE |
|||
Cultivar |
|||
| Watering regime | Rafiki |
Indiana |
Essoumtem |
| Control | 0.21±0.06b |
0.15±0.02b |
0.19±0.02b |
| Normal | 0.28±0.04b |
0.19±0.03b |
0.25±0.02b |
| Moderate | 0.37±0.12b |
0.24±0.07b |
0.25±0.03b |
| Severe | 0.65±0.09a |
0.33±0.18b |
0.28±2.45b |
| Normal+virus | 0.22±0.03b |
0.28±0.07b |
0.14±0.04b |
| Moderate+virus | 0.29±0.04b |
0.27±0.00b |
0.16±0.02b |
| Severe+virus | 0.35±0.05b |
0.30±0.14b |
0.36±0.15b |
| Means that do not share a letter are significantly different at a 95% confidence interval | |||
DISCUSSION
In this investigation, reduced plant height was recorded in the severe drought in all three okra cultivars. These cultivars were compared to their respective controls and these findings agree with similar research by Ewetola and Fasnami16, who reported plant height reduction in okra plants subjected to 25% field capacity of water as compared to plants under 75% field capacity of water. Current results also agreed with Wu et al.17, who found out that plant height in water-stressed citrus seedlings was reduced up to 25%. A decrease in plant height in drought-stressed plants is usually associated with a decline in cell enlargement and an increase in leaf senescence18. A likely explanation for reduced plant height may be that cell turgor is decreased under drought conditions leading to low cell metabolism which results in the inhibition of cell division and elongation19. In a related finding, the current study results seem to agree with reports in some wheat genotypes where water stress was shown to inhibit growth and negatively affected plant height20,21.
Virus-infected plants showed a reduction in plant height in three cultivars. The percentage reduction in plant height was pronounced in virus and severe drought treatment. Again results in this work showed percentage reduction was high in that Rafiki and Indiana but low in Essoumtem, indicating a possible tolerance to both virus and drought combination stress. This was in line with the findings of Khaskheli et al.22, who reported a significant reduction in plant height of okra grown in four different locations and infected with the yellow vein mosaic virus as compared to healthy plants. Amiteye et al.23 also reported that plant height was significantly reduced in diseased plants of all ten okra cultivars infected with the okra mosaic virus (OkMV). Essoumtem cultivar was tolerant to the virus infection as plant height reduction was insignificant when compared to the control. Similar observations were made by Salaudeen et al.24, who reported insignificant differences in plant height between uninoculated okra plants and okra plants infected with the Cassava mosaic virus (CMV) in Ex-Bassawa-2, Cayenne and Roma Savanna cultivars in a screen house experiment.
Although drought may increase the susceptibility of plants to diseases, it can, on the contrary, mitigate the negative impact of causing increased diseases in plants25. The higher the drought stress level, the higher the impact of both stresses on the plant. In this study, simultaneous stress (a combination of severe watering and virus infection) caused the highest plant height reductions in all three cultivars when compared to other treatments. In another related finding, sweet corn plants (Zea mays var. saccharata) simultaneously exposed to Maize dwarf mosaic virus and drought stress showed more reduction in plant height compared to non-infected plants26. Further, Prasch and Sonnewald27 also reported that simultaneous exposure of Arabidopsis plants to drought and Turnip mosaic virus led to a higher reduction in morphological features under combined stresses as compared to single or individual stress. Comparing all three cultivars, Indiana and Rafiki showed the highest stunted growth and thus were highly susceptible to severe doses of both stresses.
All cultivars recorded a 100% disease incidence which indicated that viral transmission through the mechanical inoculation process reflected what was observed and reported by Mahjabeen et al.28. Evidence of systemic infection was observed a week after inoculation in newly emerged leaves agreeing with the findings of Murphy et al.29, who reported similar observations on bell peppers infected with the Cucumber mosaic virus. Symptoms such as mosaic patches, yellowing of leaf vein, yellowing of leaves (chlorosis), necrosis and stunted growth observed in all cultivars in this research were similar to symptoms seen in leaves of okra plants infected by okra yellow vein mosaic virus (OYVMV)30. Stephan et al.31 also reported symptoms of okra mosaic virus infection which included mosaic patches on leaf blades, vein chlorosis, vein-banding, plant stunting and yellowing of the leaves in extreme cases as observed in this study. Since OYVMV is a begomovirus and might not be sap transmissible, there is a high possibility of OkMV being responsible for the symptoms observed in this work.
With the exception of control plants that did not show any yellowing of leaves, chlorotic symptoms observed with developmental aging of plants in this study, were found to be linked with both reduced intensity of green chlorophyll pigment and leaf area32. Doumayrou33 also mentioned that the measures of mortality in Brassica rapa and Arabidopsis thaliana were found to correlate with the severity of visual qualitative symptoms in Cauliflower mosaic virus (CaMV).
Disease symptoms recovery, observed in this work, corresponded to the findings of Nie34, who found that tobacco plants infected with the Tobacco mosaic virus go through incubation, rapid symptom progression and partial recovery stages. Viral disease recovery is usually associated with systemic acquired resistance which is the potential of plants to induce local and systemic defence responses after an initial infection35. Jovel et al.36 also reported that the Tomato Ringspot Virus (ToRSV) has been shown to use interfering RNAs and PR1a transcripts as a defence response mechanism in tomato plants. Virus recovery from plants infected with isolates of particular viruses such as the African Cassava Mosaic Virus (ACMV-[CM]) or Sri Lankan Cassava Mosaic Virus occurs because of an innate gathering of siRNA in plants inoculated with virus isolates compared to virus-infected plants without the ability to recover from symptoms37.
In this investigation, a combined effect of virus infection and drought (severe watering and virus) showed an increase in symptom severity score in all three cultivars as compared to a single effect of virus infection only. These observations appear to agree with Paudel and Sanfaçon32, who reported similar findings that external factors such as temperature, photoperiod, plants' ability to access water and carbon dioxide presence may affect the viral symptomatology. Similarly, Bergès et al.38 reported that heat, drought or salt stress enhance plant susceptibility to pathogens whereas on the other hand, viruses seem to enhance the ability of plants to counteract these abiotic stresses by inducing drought or cold tolerance.
The observation in this study revealed that drought stress caused a significant decrease in a number of leaves in all three cultivars used in this study as compared to their respective control plants. Essoumtem cultivars appear to have the ability to resist the phenomenon of leaf senescence as compared to Rafiki and Indiana cultivars. These findings were supported by Murphy et al.29, who observed that leaf numbers and their cumulative weight, both along the main stem and among branches, were significantly less for plants infected with Cassava mosaic virus (CMV). Similar findings were reported by Pazarlar et al.39 in a study to determine the effects of Tobacco mosaic virus infection on growth and physiological parameters in some pepper varieties (Capsicum annuum L.) These findings appear to be similar to reports by Kusvuran40, who reported that drought stress decreased the leaf number of okra genotypes. In addition, findings in this work corresponded to reports by Nguyen et al.41, who mentioned that under severe water deficit, there was partial or complete loss of leaves in four dominant tree species of the Asian savanna. On the contrary, investigations by Al-Ubaydi et al.42 agreed with the results of this work that watering okra plants every three days significantly gave a higher number of leaves as compared with delayed watering over a long period. Possible reasons for the observed phenomenon may be that water stress causes a decrease in the initiation of new leaves and increases the rate of leaf fall. Essoumtem cultivar however showed tolerance traits as the number of leaves in virus-infected plants did not vary significantly from control plants.
All three cultivars exposed to simultaneous drought and virus infection had significantly reduced the number of leaves when compared to control plants. These observations were similar to findings reported in Giró Ros plants43. It appears that Rafiki and Essoumtem activated basal defence responses when exposed to mild drought and virus stresses which enabled these plant cultivars to defend against the pathogen infection in the combined virus and severe drought infection. On the contrary, it has been shown that severe drought causes leakage of cellular nutrients into the apoplast which facilitates successful pathogen infection13. Cultivars used in this study which were treated under combined severe and virus stresses exhibited susceptibility to pathogen infection as compared to single virus stress.
In this study, internode length decreased with increasing drought severity. These observations have also been reported by Bayer et al.44 in Hibiscus acetosella ‘Panama Red’ grown with a soil moisture sensor-controlled irrigation system. In this study, reduced internode lengths were observed in all three cultivars under severe drought as compared to their respective control plants. Reduction in stem elongation under drought has been associated with shorter internode lengths45. It has also been established that stem elongation is facilitated by cell division and expansion and it is a common marker of drought stress responses in most plants46. Interestingly, reductions in stem elongation could be a result of acclimation and sometimes increases with increasing stress47. Feng et al.48 in a study on soybean mentioned that limited water supply reduced average internode length by 33% maturity. In another experiment to screen for drought tolerant pear populations, Tatari et al.49 reported that drought stress led to a reduction in internode length in all pear populations. Similar observations were also reported by Reyes et al.50.
Virus-infected plants had significantly shorter internode lengths in all three cultivars when compared to their controls and even when compared to plants under all drought stress levels. Osmani et al.51 reported similar findings where morphological responses of potato plants to virus stresses showed a significant decline in plant growth-related traits such as stem and internode length, stem diameter and leaf size as compared to healthy control plants. In a related study, Murphy et al.29 observed a significant reduction in internode extension of Cucumber mosaic virus (CMV) infected plants when compared to internode length of healthy control plants.
All three cultivars under a combination of both virus infection and severe drought stress had a reduced internode length when compared to either virus or drought stress as well as controls which had no virus infection or drought. The effect was pronounced in Indiana cultivar even when compared among all other treatments. In Rafiki and Essoumtem cultivars, the higher the severity of the stress combination, the shorter the internode length even though they were not significantly different. This observation suggested that different drought stress levels when combined with virus infection caused similar changes in internode lengths in Rafiki and Essoumtem cultivars.
Stem diameter reduction appear significantly reduced in Indiana cultivar than Rafiki and Essoumtem when subjected to sever water deficit conditions in this study. Kusvuran40 reported a decrease in stem diameter in drought stressed okra plants, a phenomenon, which were in agreement with results from this study. Findings in this study also showed that drought stress reduced stem diameter in all three cultivars and these revelations were similar to that made by Misra et al.52, who observed a 7.37% decrease in stalk diameter in unrelated crop such as sugarcane under drought conditions. Similarly, Reyes et al.50 also observed significantly higher stem diameter in well-watered sugarcane plants as compared to those grown under drought stress. About 22.7% reduction in stem diameter was observed in two Barbados cherry genotypes under 25% of field capacity of water as compared to control plants53. Similar findings were also reported at the reproductive growth stage in maize by Sabiel et al.54.
Stem girth in virus infected okra plants has been shown to significantly decreases55. Stem diameter in Rafiki and Indiana cultivars infected with the virus was reduced when compared to healthy control plants and this observation agreed with Wang et al.56, who reported a significant increase in stem diameter by 0.52 cm in virus free chewing cane seedlings as compared to infected ones. On the contrary, Essoumtem, recorded high stem diameter in virus infected plants when compared to healthy control plants even though the difference was not significant. Similar findings were reported by Pazarlar et al.39, where stem diameter increased by 2.12% in a Tobacco mosaic virus infected pepper variety (Kamsal) as compared to control plants.
Stem diameter was decreased in all three cultivars when compared to control plants and plants under all other treatments. Thus, simultaneous stress conditions had negative effects on plant growth and development causing significant reductions in stem diameter.
Leaf area in all three cultivars under severe drought stress reduced significantly when compared to their respective controls. This finding was supported by several studies including that of Dong et al.57, who reported that drought stress inhibited the growth of the soybean plants in terms of height and leaf area. Water deficit leads to a decrease in leaf area which is caused by inhibition of leaf expansion due to decreased rate of cell division58. A decline in leaf area results in decreased water loss due to low transpiration rates and this has been shown to be a drought avoidance strategy used by plants under drought stress5. Other authors have shown similar correlation between severe drought stress and a reduction in leaf area52,53,59. Plants generally show reduction in leaf area as a strategy to adapt to drought stress. The decrease in leaf area is as a result of decrease in leaf turgor pressure and the rate of photosynthesis of plant leaves60.
Virus infection caused a reduction in leaf area in all three cultivars and this finding is similar to that of Adeniji et al.61, who reported that leaf area per plant at 10 weeks after virus inoculation was higher in uninoculated plants compared to the inoculated plants for both genotypes. Telfairia mosaic virus infection caused significant reduction in leaf area of Sphenostylis stenocarpa62. Appiah et al.63 also reported reduction in leaf size of groundnut plants affected with the groundnut rosette disease.
In a study to evaluate the impact of drought stress and simultaneously occurring pathogen infection in field-grown chickpea, Sinha et al.26 observed a decrease in specific leaf area (SLA) in plants growing under this combined stress, indicating the negative impact of combined stresses on plant growth.
Root to shoot ratio increased in all three cultivars grown under severe watering conditions, especially Rafiki cultivar when compared with their respective controls. This finding was also supported by Makbul et al.64, who reported root to shoot ratios of 1.51 and 2.04 in plants under drought stress and unstressed plants respectively. In a similar study carried out by Ayeh et al.65 on four cowpea genotypes, high root to shoot ratios were observed in drought stressed plants as compared to their controls. Moreover, it was observed that a decrease in water availability led to an increase in root length in cultivars used in the study by Ayeh et al.65. On the contrary, Feng et al.48 reported that water stress significantly reduced the total root length and root surface area in soybean. It has been established that, when there is accumulation of roots (root length density) with corresponding increase in root diameter, drought tolerance prevails as root grows deep into the soil to obtain water whereas on the contrary, some plants show decreased root length diameter to indicate drought susceptibility26.
Virus infection caused a reduction in root to shoot ratio in Essoumtem cultivar and an increase in the same parameter in Rafiki and Indiana cultivars. However, these differences observed were not significantly different from their respective controls. This showed that shoot growth was high than root growth in Essoumtem as compared to the control, indicating that virus infection increased shoot growth. It also showed a possible resistance of Essoumtem cultivar and susceptibility of Indiana and Rafiki cultivars to virus infection.
The combination of virus infection and drought resulted in high root to shoot ratios and decreased root length as compared to other treatments. Similarly, Sinha et al.26 reported that chickpea plants subjected to combined drought with Fusarium solani on one hand and drought combined with Rhizoctonia bataticola in a separate experiment showed decreased primary root length as compared to plants treated with only pathogen infection. In another research to support findings in this research, Sheikh55 reported a 15.5% decrease in the root length of okra infected with Bhendi yellow vein mosaic virus (BYVMV) was observed.
The implication of the study is that both biotic and abiotic stress factors cause different symptoms in plants. When plants are exposed to a combination of both virus and drought stress factors, the symptoms showed a severer form of the symptoms imposed by the individual biotic and drought stress factors. The study has relevant applications in the field of plant stress biology. This research provides a knowledge and understanding of how plants behave in the open field when faced with a complexity of biotic and abiotic stress factors. However, the limitations of the studies are that it fails to reveal a clear distinction between symptoms imposed by either a virus effect or those caused by a drought effect. This distinction could have been since introduction of molecular and serological methods such as Polymerase Chain Reaction (PCR) and Enzyme Linked Immunoabsorbent Assay (ELISA) respectively to identify specific viruses infecting Okra plants and those symptoms brought about by drought stress only. Both PCR and ELISA methods were, however, beyond the scope of this study.
CONCLUSION
The current study found that virus infection and drought-stressed plants may either result in impaired growth in okra cultivars used in this study. Varying virus symptoms were observed in all three okra cultivars. Symptoms included mosaic patches on leaves yellowing of leaf veins, yellowing of leaves and clearing of leaf veins. Indiana cultivar recorded the highest symptom severity scores and can be described as the most susceptible cultivar to virus infection with Essoumtem being the most tolerant cultivar. Both stresses caused reductions in all morphological traits investigated in this study with a combination of both stresses being the most damaging. Severe drought stress caused significant morphological impacts on plant height, number of leaves, stem diameter and leaf area. Thus drought stress caused an increased reduction in growth as compared to virus-infected plants. However, the contrary was observed in internode length and root to shoot ratio.
SIGNIFICANCE STATEMENT
This study discovered that the combined stress of drought and virus pathogen had a profound effect on plant growth and development and these responses to drought and viruses were in some cases cultivar specific. Further, the study gave insights into the potential effect of drought, virus and combined drought and virus as tolerance screening strategies in okra plants. The study will be beneficial to the world of plant sciences because the subject of combined stress effects on plants is rarely studied in the laboratory and this underscores the need to study individual and combined stresses effect on plants which may mimic field conditions.
ACKNOWLEDGMENTS
Gratitude goes to the technical staff, Department of Plant and Environmental Biology, University of Ghana and Biotechnology and Nuclear Agriculture Institute of Ghana, Kwabenya for the technical assistance that saw to the success of this research.
REFERENCES
- Mishra, G.P., B. Singh, T. Seth, A.K. Singh and J. Halder et al., 2017. Biotechnological advancements and begomovirus management in Okra (Abelmoschus esculentus L.): Status and perspectives. Front. Plant Sci., 8: 00360.
- Schafleitner, R., C.Y. Lin, Y.P. Lin, T.H. Wu and C.H. Hung et al., 2021. The world vegetable center Okra (Abelmoschus esculentus) core collection as a source for flooding stress tolerance traits for breeding. Agriculture, 11.
- Vwioko, E.D., M.A. El-Esawi, M.E. Imoni, A.A. Al-Ghamdi and H.M. Ali et al., 2019. Sodium azide priming enhances water logging stress tolerance in Okra (Abelmoschus esculentus L.). Agronomy, 9.
- Singha, P., V. Chauhana, B.K. Tiwaria, S.S. Chauhanb, S. Simonb, S. Bilalc and A.B. Abidia, 2014. An overview on okra (Abelmoschus esculentus) and its importance as a nutritive vegetable in the world. Int. J. Pharm. Bio. Sci., 4: 227-233.
- Havrlentová, M., J. Kraic, V. Gregusová and B. Kovácsová, 2021. Drought stress in cereals-A review. Agriculture (Pol'nohospodárstvo), 67: 47-60.
- Gursoy, M., A. Balkan and H. Ulukan, 2012. Ecophysiological responses to stresses in plants: A general approach. Pak. J. Biol. Sci., 15: 506-516.
- Singla, J. and S.G. Krattinger, 2016. Biotic Stress Resistance Genes in Wheat. In: Encyclopedia of Food Grains, Wrigley, C., H. Corke, K. Seetharaman and J. Faubion (Eds.), Elsevier Academic Press, USA, ISBN: 978-0-12-394786-4, pp: 388-392.
- Encabo, J.R., R.J.A. Macalalad-Cabral, J.M.K. Matres, S.C.T.P. Coronejo and G.B. Jonson et al., 2020. Infection with an asymptomatic virus in rice results in a delayed drought response. Funct. Plant Biol., 47: 239-249.
- Rivero, R.M., T.C. Mestre, R. Mittler, F. Rubio, F. Garcia-Sanchez and V. Martinez, 2014. The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environ., 37: 1059-1073.
- Gupta, A., S.K. Dixit and M. Senthil-Kumar, 2016. Drought stress predominantly endures Arabidopsis thaliana to Pseudomonas syringae infection. Front. Plant Sci., 7.
- Dahivelkar, P., G.E. Atre, P.V. Gawande and G.D. Mate, 2017. Management of powdery mildew of Okra caused by Erysipheci choracearum., Int. J. Curr. Microbiol. Appl. Sci., 6: 3189-3193.
- Pandey, P., V. Irulappan, M.V. Bagavathiannan and M. Senthil-Kumar, 2017. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front. Plant Sci., 8.
- Ramegowda, V. and M. Senthil-Kumar, 2015. The interactive effects of simultaneous biotic and abiotic stresses on plants: Mechanistic understanding from drought and pathogen combination. J. Plant Physiol., 176: 47-54.
- van Munster, M., M. Yvon, D. Vile, B. Dader, A. Fereres and S. Blanc, 2017. Water deficit enhances the transmission of plant viruses by insect vectors. PLoS ONE, 12: 0174398.
- Musa, U. Tanko and T.H. Usman, 2016. Leaf area determination for maize (Zea mays L.), Okra (Abelmoschus esculentus L.) and cowpea (Vigna unguiculata L.) crops using linear measurements. J. Biol. Agric. Healthcare, 6: 103-111.
- Ewetola, E.A. and T.F. Fasanmi, 2022. Growth responses of Okra (Albemoschus esculentus) and jute mallow (Corchorus oitorius) to water stress and non-water stress conditions. Int. Lett. Chem, Phys. Astron., 59: 10-16.
- Wu, Q.S., R.X. Xia and Y.N. Zou, 2008. Improved soil structure and citrus growth after inoculation with three arbuscular mycorrhizal fungi under drought stress. Eur. J. Soil Biol., 44: 122-128.
- Begna, T., 2020. Effects of drought stress on crop production and productivity. Int. J. Res. Stud. Agric. Sci., 6: 34-43.
- Patmi, Y.S., A. Pitoyo, Solichatun and Sutarno, 2020. Effect of drought stress on morphological, anatomical, and physiological characteristics of Cempo Ireng cultivar mutant rice (Oryza sativa L.) strain 51 irradiated by gamma-ray. J. Phys. Conf. Ser., 1436.
- Sun, S., L. Liu and S. Yang, 2015. Research on the effects of water stress on growth traits and water use efficiency of winter wheat. MATEC Web Conf., 25: 01010.
- Pour-Aboughadareh, A., R. Mohammadi, A. Etminan, L. Shooshtari, N. Maleki-Tabrizi and P. Poczai, 2020. Effects of drought stress on some agronomic and morpho-physiological traits in durum wheat genotypes. Sustainability, 12: 5610.
- Khaskheli, M.I., M.M. Jiskani, S.P. Goswami, G.B. Poussio and M.A. Khaskheli, 2017. Effect of okra yellow vein mosaic virus (OYVMV) on plant growth and yield. J. Basic Appl. Sci., 13.
- Amiteye, S., A.S. Appiah, F. Boateng, J.T. Kutufam and H.M. Amoatey, 2021. Physiological changes associated with Okra mosaic virus infection in field grown okra plants. Asian J. Agric. Biol., 2021: 4.
- Salaudeen, M.T., O. Oluwatosin and A.S. Gana, 2018. Reactions of commercial cultivars of okra, pepper, and tomato to cucumber mosaic virus disease. Agro-Science, 17: 27-36
- Hosseini, S.A., G.R. Zamani, Z.M. Yaghub and M. Khayyat, 2018. Effects of Cucumber Mosaic Virus infection and drought tolerance of tomato plants under greenhouse conditions: Preliminary results. J. Berry Res., 8: 129-136
- Sinha, R., V. Irulappan, B. Mohan-Raju, A. Suganthi and M. Senthil-Kumar, 2019. Impact of drought stress on simultaneously occurring pathogen infection in field-grown chickpea. Sci. Rep., 9: 5577.
- Prasch, C.M. and U. Sonnewald, 2013. Simultaneous application of heat, drought, and virus to Arabidopsis plants reveals significant shifts in signaling networks. Plant Physiol., 162: 1849-1866
- Mahjabeen, K.P. Akhtar, N. Sarwar, M.Y. Saleem, M. Asghar, Q. Iqbal and F.F. Jamil, 2012. Effect of cucumber mosaic virus infection on morphology, yield and phenolic contents of tomato. Arch. Phytopathol. Plant Prot., 45: 766-782
- Murphy, J.F., J. Masiri, B.A. R. Hadi and R.R. Dute, 2016. Recovery from Cucumber mosaic virus infection for ‘Calwonder’ bell pepper plants does not counter negative impacts on plant growth. J. Phytopathol. 164: 840-846
- Senevirathna, H.M.S.I., S.K. Wasala, D.M.J.B. Senanayake, D. Weerasekara, H.A.M. Wickamasinghe and P.K.G.A. Deepal, 2016. Characterization and detection of yellow vein disease of Okra (Abelmoschus esculentus (L.) moench) in Sri Lanka. Trop. Agric. Res., 27: 360-369
- Stephan, D., M. Siddiqua, A.T. Hoang, J. Engelmann, S. Winter and E. Maiss, 2008. Complete nucleotide sequence and experimental host range of Okra mosaic virus. Virus Genes, 36: 231-240
- Paudel, D.B. and H. Sanfaçon, 2018. Exploring the diversity of mechanisms associated with plant tolerance to virus infection. Front. Plant Sci., 9. https://doi.org/10.3389/fpls.2018.01575
- Doumayrou, J., S. Leblaye, R. Froissart and Y. Michalakis, 2013. Reduction of leaf area and symptom severity as proxies of disease-induced plant mortality: The example of the Cauliflower mosaic virus infecting two Brassicaceae hosts. Virus Res., 176: 91-100
- Nie, X., 2006. Salicylic acid suppresses potato virus Y isolate N:O-induced symptoms in tobacco plants. Phytopathology, 96: 255-263
- Stahl, E., P. Bellwon, S. Huber, K. Schlaeppi and F. Bernsdorff et al., 2016. Regulatory and functional aspects of indolic metabolism in plant systemic acquired resistance. Mol. Plant, 9: 662-681
- Jovel, J., M. Walker and H. Sanfaçon, 2011. Salicylic acid-dependent restriction of Tomato ringspot virus spread in tobacco is accompanied by a hypersensitive response, local RNA silencing, and moderate systemic resistance. Mol. Plant-Microbe Interact., 24: 706-718
- Chellappan, P., R. Vanitharani and C.M. Fauquet, 2004. Short interfering RNA accumulation correlates with host recovery in DNA virus-infected hosts, and gene silencing targets specific viral sequences. J. Virol., 78: 7465-7477
- Bergès, S.E., D. Vile, C. Vazquez-Rovere, S. Blanc and M. Yvon et al., 2018. Interactions between drought and plant genotype change epidemiological traits of cauliflower mosaic virus. Front. Plant Sci., 9: .
- Pazarlar, S., M. Gumus and G.B. Oztekin, 2013. The effects of Tobacco mosaic virus infection on growth and physiological parameters in some pepper varieties (Capsicum annuum L.). Not. Bot. Hortic. Agrobotanici Cluj-Napoca, 41: 427-433.
- Kusvuran, S., 2012. Influence of drought stress on growth, ion accumulation and antioxidative enzymes in okra genotypes. Int. J. Agric. Biol., 14: 401-406.
- Nguyen, T.T., S.K. Arndt and P.J. Baker, 2019. Leaf physiological responses to drought stress and community assembly in an Asian Savanna. Forests, 10.
- Al-Ubaydi R.M., F.E. Al-Shakry, A.M. Al-Samara and M.S. Al-Mohmadawy, 2017. Effect of irrigation intervals on growth, flowering and fruits quality of Okra Abelmoschus esculentus (L.) monech. Afr. J. Agric. Res., 12: 2036-2040.
- El Aou-ouad, H., J. Bota, T. Obata, R. Montero, A.R. Fernie, H. Medrano, A. Pou and I. Florez-Sarasa, 2018. Combined drought and virus infection trigger aspects of respiratory metabolism related to grapevine physiological responses. J. Plant Physiol., 231: 19-30.
- Bayer, A., I. Mahbub, M. Chappell, J. Ruter and M.W. van Iersel, 2013. Water use and growth of Hibiscus acetosella ‘panama red’ grown with a soil moisture sensor-controlled irrigation system. HortScience, 48: 980-987.
- Morales, R.G.F., L.V. Resende, I.C. Bordini, A.G. Galvão and F.C. Rezende, 2015. Characterization of tomato plants submitted to water deficit [In Portuguese]. Sci. Agraria, 16: 9-17.
- Avramova, V., H. AbdElgawad, Z. Zhang, B. Fotschki and R. Casadevall et al., 2015. Drought induces distinct growth response, protection, and recovery mechanisms in the maize leaf growth zone. Plant Physiol., 169: 1382-1396.
- Litvin, A.G., M.W. van Iersel and A. Malladi, 2016. Drought stress reduces stem elongation and alters gibberellin-related gene expression during vegetative growth of tomato. J. Am. Soc. Hortic. Sci., 141: 591-597.
- Feng, Y.Y., J. He, N.C. Turner, K.H.M. Siddique and F.M. Li, 2021. Phosphorus supply increases internode length and leaf characteristics, and increases dry matter accumulation and seed yield in soybean under water deficit. Agronomy, 11: 11050930.
- Tatari, M., A. Jafari, M. Shirmardi and M. Mohamadi, 2019. Using morphological and physiological traits to evaluate drought tolerance of pear populations (Pyrus spp.). Int. J. Fruit Sci., 20: 837-854.
- Reyes, J.A.O., A.S. Carpentero, P.J.A. Santos and E.F. Delfin, 2020. Effects of water regime, genotype, and formative stages on the agro-physiological response of sugarcane (Saccharum officinarum L.) to drought. Plants, 9.
- Osmani, Z., M.S. Sabet, K.S. Nakahara, A. Mokhtassi-Bidgoli, K. Vahabi, A. Moieni and M. Shams-Bakhsh, 2020. Identification of a defense response gene involved in signaling pathways against PVA and PVY in potato. GM Crops Food, 12: 86-105.
- Misra, V., S. Solomon, A.K. Mall, C.P. Prajapati and A. Hashem et al., 2020. Morphological assessment of water stressed sugarcane: a comparison of waterlogged and drought affected crop. Saudi J. Biol. Sci., 27: 1228-1236.
- Medeiros, D.B., E.C. da Silva, H.R.B. Santos, C.M. Pacheco, R. dos Santos Musser and R.J.M.C. Nogueira, 2013. Physiological and biochemical responses to drought stress in Barbados cherry. Braz. J. Plant Physiol., 24: 181-192.
- Sabiel, S.A.I., A.A. Abdelmula, E.M.A. Bashir, S. Khan and S. Yingying et al., 2013. Genetic variation of plant height and stem diameter traits in maize (Zea mays L.) under drought stress at different growth stages. J. Nat. Sci. Res., 4: 116-122.
- Sheikh, M.A., Safiuddin, Z. Khan and I. Mahmood, 2013. Effect of bhendi yellow vein mosaic virus on yield components of okra plants. J. Plant Pathol., 95: 391-393.
- Wang, K.L., Q.Q. Deng, J.W. Chen and W.K. Shen, 2020. Physiological and molecular mechanisms governing the effect of virus-free chewing cane seedlings on yield and quality. Sci. Rep., 10.
- Dong, S., Y. Jiang, Y. Dong, L. Wang and W. Wang et al., 2019. A study on soybean responses to drought stress and rehydration. Saudi J. Bio. Sci., 26: 2006-2017.
- Kapoor, D., S. Bhardwaj, M. Landi, A. Sharma, M. Ramakrishnan and A. Sharma, 2020. The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Appl. Sci., 10.
- Petrović, I., Z. Jovanović, R. Stikić, M. Marjanović and S. Savić, 2021. Influence of severe drought on leaf response in aba contrasting tomato genotypes (wild type and flacca mutant). Biol. Life Sci., 4.
- Yang, X., M. Lu, Y. Wang, Y. Wang, Z. Liu and S. Chen, 2021. Response mechanism of plants to drought stress. Horticulturae, 7.
- Adeniji, M.O., S.A. Shoyinka, T. Ikotun, R. Asiedu, J.d'A Hughes and B.O. Odu, 2012. Yield loss in Guinea yam (Dioscorea rotundata poir.) due to infection by Yam mosaic virus (YMV) genus Potyvirus. Ife J. Sci., 14: 237-244.
- Mofunanya, A.A.J., V.B. Ogar, J.O. Oni, T.E. Omarachong and F.A. Akomaye, 2020. Growth and yield assessment of Sphenostylis stenocarpa affected by virus infection. Iran. J. Plant Physiol., 11: 3433-3441.
- Appiah, A.S., S.K. Offei, R.S. Tegg and C.R. Wilson, 2016. Varietal response to groundnut rosette disease and the first report of Groundnut ringspot virus in Ghana. Plant Dis., 100: 946-952.
- Makbul, S., N.S. Guler, N. Durmus and S. Guven, 2011. Changes in anatomical and physiological parameters of soybean under drought stress. Turk. J. Bot., 35: 369-377.
- Ayeh, K.O., A.K. Peter, A.E. Grace and L. Enu-Kwesi, 2021. Physiological, developmental and growth responses to desiccation induced stress in four seed coat colour varieties of Vigna unguiculata (L. Walp). Afr. J. Agric. Res., 17: 642-657.
How to Cite this paper?
APA-7 Style
Amadu,
A.M., Appiah,
A.S., Ayeh,
K.O. (2023). Differential Biotic and Abiotic Induced Stress Levels Confer Varying Morphological Responses in Okra (Abelmoschus esculentus L. Moench) Cultivars. Asian Journal of Biological Sciences, 16(3), 302-321. https://doi.org/10.3923/ajbs.2023.302.321
ACS Style
Amadu,
A.M.; Appiah,
A.S.; Ayeh,
K.O. Differential Biotic and Abiotic Induced Stress Levels Confer Varying Morphological Responses in Okra (Abelmoschus esculentus L. Moench) Cultivars. Asian J. Biol. Sci 2023, 16, 302-321. https://doi.org/10.3923/ajbs.2023.302.321
AMA Style
Amadu
AM, Appiah
AS, Ayeh
KO. Differential Biotic and Abiotic Induced Stress Levels Confer Varying Morphological Responses in Okra (Abelmoschus esculentus L. Moench) Cultivars. Asian Journal of Biological Sciences. 2023; 16(3): 302-321. https://doi.org/10.3923/ajbs.2023.302.321
Chicago/Turabian Style
Amadu, Ayisha, Marfo, Andrew Sarkodie Appiah, and Kwadwo Owusu Ayeh.
2023. "Differential Biotic and Abiotic Induced Stress Levels Confer Varying Morphological Responses in Okra (Abelmoschus esculentus L. Moench) Cultivars" Asian Journal of Biological Sciences 16, no. 3: 302-321. https://doi.org/10.3923/ajbs.2023.302.321

This work is licensed under a Creative Commons Attribution 4.0 International License.