Morphology and Agronomic Evaluation of Ten Mungbean (Vigna radiata L.) Accessions in Derived Savanna Agroecology of Southeast Nigeria
-
Uchenna Noble Ukwu


Department of Crop Science, Faculty of Agriculture, University of Nigeria, 410105, Nsukka, Enugu, Nigeria
Blessing Ngozika OburuDepartment of Crop Science, Faculty of Agriculture, University of Nigeria, 410105, Nsukka, Enugu, Nigeria
Stella Ogochukwu MuojiamaDepartment of Crop Science and Horticulture, Nnamdi Azikiwe University, 420110, Awka, Nigeria
Vivian Ogechi OsadebeDepartment of Crop Science and Horticulture, Nnamdi Azikiwe University, 420110, Awka, Nigeria
Nathaniel DaudaDepartment of Crop Science, Faculty of Agriculture, University of Nigeria, 410105, Nsukka, Enugu, Nigeria
Solomon Oluwaseyi AdewuyiDepartment of Crop Science, Faculty of Agriculture, University of Nigeria, 410105, Nsukka, Enugu, Nigeria
| Received 04 Jan, 2023 |
Accepted 10 Apr, 2023 |
Published 30 Jun, 2023 |
Background and Objective: Mungbean is relatively unknown and underutilized pulse in Southeast Nigeria with enormous potential to eliminate hunger and malnutrition in the region. This study was carried out to evaluate, characterize and select promising mungbean genotypes based on morphological and agronomic traits in an attempt to increase the legume base and secure alternative sources of plant-based proteins for Nigerians. Materials and Methods: Ten mungbean accessions were characterized and evaluated between April to October, 2020 at the Department of Crop Science, University of Nigeria, in a randomized complete block design (RCBD) with four replications. Data were collected on 23 morphological and 21 agronomic traits. Results: The clear-cut variation in 21 of 23 morphological traits and 16 of 21 agronomic traits. The number of pods (NOP) per plant, number of seeds (NOS) per pod and seed yield were the most discriminating yield indices. The Tvr18 and Tvr79 recorded higher NOP per plant, NOS per pod and seed yield and were therefore selected for advancement to the next generation. The ten accessions were clustered into two distinct groups, cluster I and II, with 8 and 2 accessions, respectively. The Tvr83 showed uniqueness for a higher number of leaves (NOL), number of branches (NOB), NOP per plant (although with shorter pods) and smaller leaves compared to the other nine accessions. Conclusion: The two high yielding accessions, Tvr18 and Tvr79 could provide flatulence free proteins to Nigerians while, Tvr83 is a valuable candidate for improvement of Tvr18 and Tvr79 in any of these traits for which it showed superiority.
| Copyright © 2023 Ukwu et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
By 2030, the second sustainable development objective of the UN purposes to completely exterminate hunger and food insecurity1. This goal may be unrealistic if efforts are concentrated solely on major staple food crops. In view of the predicted upsurge in the global population from 7.7 billion in 2020 to 8.5 billion in 20302, there is an increasing need to explore other minor or less known crops. Mungbean is a minor pulse in Southeast Nigeria since its first introduction in 20033. Mungbean improvement programs have been limited due to low genetic diversity among the mungbean gene pool4,5. Consequently, statistical data for other pulses are not readily available globally in FAO statistical database due to scanty research on the crop6,7. Mungbean yields in many countries are considerably “low, ranging from 0.5-1.5 t ha–1”7. The Asian Vegetable Research and Development Centre (AVRDC) had made several attempts to promote mungbean research globally through seed distribution among countries to deliver superior germplasm with increased reliability and profitability for growers7. In spite of AVRDC’s efforts, mungbean research in many countries is still poorly funded due to its minor status and the poor awareness of the enormous nutritional, medicinal and environmental benefits of the crop. Previous evaluation studies across agroecological zones in Nigeria have shown mungbean to be a low-cost crop, highly nutritious but low in yield3,8-12. In the derived savanna zone of Southeast Nigeria, there is scarce literature report on mungbean evaluation studies. Sufficient yield variability could exist among mungbean germplasm and successful identification of high-yielding genotypes could be pivotal in the struggle to eliminate hunger by 2030 as mungbean could provide a cheap alternative source of non-flatulence protein13, essential amino acids such as phenylalanine, leucine, isoleucine, lysine, arginine14, vitamins, minerals, fiber, resistant starch and antioxidants which decreases the risk of chronic diseases15. In addition, it improves soil fertility through its role in nitrogen fixation and environmental conservation12,16,17 and serves as an integral part of therapeutic medicine18. The objective of this study was to evaluate, characterize and select promising mungbean genotypes based on morphological, agronomic and yield traits.
MATERIALS AND METHODS
Two field experiments were carried out at the Experimental Field of the Department of Crop Science, University of Nigeria, Nsukka (latitude 06°52'N, longitude 07°24'E and altitude 447) between, April and October, 2020.
Ten mungbean genotypes (Tvr18, Tvr19, Tvr24, Tvr28, Tvr32, Tvr34, Tvr49, Tvr65, Tvr79 and Tvr83) sourced from the Genetic Resource Centre, International Institute of Tropical Agriculture (IITA), Ibadan (Table 1) were replicated four times in RCBD. Plants were spaced 30×10 cm apart within plots. Each 0.6 m2 plot size contained 20 stands with a 1 m gap between plots. Land preparation, seed priming, seeding, manure application, thinning and weed management were carried out following the standard procedures used by Ihejiofor et al.12.
Data were collected on 23 morphological descriptors and 16 agronomic parameters according to the standard procedures used by Kaur et al.19. The number of days (NOD) to seed emergence was determined by recording the NOD from the day the seeds were sown till the day of seed emergence. The NOD to first and 50% flowering were obtained by recording the days from the date of seeding to when at least one and half of the plant flower, respectively. The NOD to pod formation was recorded by calculating the days from seeding to when at least the first pod is formed.
| Table 1: | Passport data of ten mungbean accessions evaluated in derived savanna agro-ecology of Southeast Nigeria | |||
| Accession number | Specie name |
Donor’s name |
Donor’s number |
Local name |
Collection date |
|
| Tvr18 | Vigna radiata L. |
IITA |
00BE14 |
Mungbean |
819 |
|
| Tvr19 | ” |
” |
” |
” |
” |
|
| Tvr24 | ” |
” |
” |
” |
” |
|
| Tvr28 | ” |
” |
” |
” |
” |
|
| Tvr32 | ” |
” |
” |
” |
” |
|
| Tvr49 | ” |
” |
” |
” |
” |
|
| Tvr65 | ” |
” |
” |
” |
” |
|
| Tvr79 | ” |
” |
” |
” |
” |
|
| Tvr83 | ” |
” |
” |
” |
” |
|
| Tvr34 | ” |
” |
” |
” |
” |
|
| IITA: International institute of tropical agriculture, Ibadan, Nigeria | ||||||
The NOL per plant was obtained by counting the number of fully expanded leaves per plant. Plant height was recorded as the height from the ground to the apex of the terminal leaf buds using tape. Leaf area was determined following the procedure of Hamid and Agata20 with measurements taken on the 4th fully expanded leaf from the apex. Stem diameter was determined with a measuring tape at 5 cm above ground level of each plant. The NOS per pod was determined by taking the average number of seeds from 10 pods per plant. Average seed weight was determined as total seed weight per plant divided by the NOS per plant. Seed length was recorded as the mean length of 10 seeds selected at random, not including those at both ends of the pods. Seed width was recorded as the mean distance between the hilum and the keel of 10 seeds. Seed thickness was recorded as the mean thickness of 10 seeds measured for length and measurement was made perpendicular to length and width. Pod length was recorded as the average length of 10 pods per plant with a measuring tape. Pod width was recorded as the average width of 10 pods measured for length by use of a DIN862 ABS Mitutoyo Digital Vernier Caliper Seed yield per plant was recorded as the total weight of dry seeds per plant.
Statistical analysis: To test for the significance of treatment means, Analysis of Variance (ANOVA) was performed on all data using Genstat 12th edition. Post-ANOVA means separation was achieved using the least significant difference (LSD) at p<0.05. Graphs were constructed using Graphpad prism 6 and dendogram was made with IBM SPSS 21.
RESULTS
Variation in phenology of ten mungbean accessions evaluated in derived savanna agro-ecology of Southeast Nigeria: The effect of genotype on the phenology of mungbean was shown in Fig. 1a. Days to first and 50% seedling emergence were similar across all ten genotypes with a mean number of 3.5 and 4.2 days after planting (DAP) to first and 50% emergence, respectively. Contrarily, the genotypes responded differently to NOD to flowering (p<0.05). Although, all genotypes exhibited asynchronous flowering habit, Tvr32, Tvr49 and Tvr83 took the least NOD to first flowering (40 DAP) and 50% flowering (42 DAP) to Tvr65 which was the latest, taking 48.33 and 53.43 DAP to first and 50% flowering, respectively (Fig. 1a). The NOD to first podding was earliest in Tvr49 (41.70), Tvr32 (42.00) and Tvr83 (42.67) in contrast to Tvr65 (54.00) and Tvr18 (50.67) which were the latest. The same trend was noticeable for NOD to 50% podding. Tvr83, Tvr24, Tvr49 and Tvr32 attained 50% podding in ≤50 DAP compared to Tvr65 and Tvr18 which took 63.65 and 66.20 DAP to attain 50% podding.
Variation in morphology of ten mungbean accessions evaluated in derived savanna agro-ecology of Southeast Nigeria: Variations in morphological attributes of ten mungbean accessions were shown in Table 2a and b. All genotypes showed indeterminate growth habits. Six were of the erect type, three semi-erect and one of the spreading type. Hypocotyl colour varied from green (2) to greenish purple (7) to purple (1). Six accessions had cuneate leaf shape, three-deltoid and one-ovate. Seven accessions had medium leaves, two-large and one-small leaves with leaf pubescence present in all accessions (Plate 1). Greenish-purple was the dominant petiole colour (7 of 10), six of which were medium in length, two were long and two were short. With an exception of Tvr83 which had its raceme below the canopy, all other nine accessions had racemes above the canopy. Calyx colour varied from greenish-purple (4) to greenish-gray (3), light-gray (1), green (1) and greenish-yellow (1). The dominant corolla colour was greenish-yellow (7) while the immature pod colour ranged from light green (8) to deep green (2). All pods were of the round type with heavily present pubescence in seven compared to three intermediates. Nine of the accessions had the least curved pods while one had medium-curved pods. Flower colour varied from pale yellow (8) to light (1) and bright yellow (1). Most accessions had mixed coloured seeds with dull luster in nine accessions while, only Tvr83 had shiny seeds (Plate 2). With an exception of Tvr83 which had oval seeds, all nine accessions had round seeds. Constrictions of pods between seeds were present in all the accessions.
| Table 2a: | Morphological characteristics of ten mungbean (Vigna radiata L.) accessions in derived savanna agro-ecology of Southeastern Nigeria | |||
| Accession | Growth habit |
Growth pattern |
Hypocotyl color |
Terminal leaf shape |
Terminal leaf length |
Leaf Pube- scence |
Leaf color |
Petiole color |
Petiole length |
Peduncle length |
Raceme position |
Calyx color |
Corolla-color |
| Tvr18 | Indeterminate |
Erect |
Greenish purple |
Cuneate |
L |
+ |
GN |
Greenish purple |
Long |
Short |
1 |
Light gray |
Greenish yellow |
| Tvr19 | Indeterminate |
Erect |
Greenish purple |
Cuneate |
M |
+ |
GN |
Greenish purple |
Medium |
Short |
1 |
Greenish purple |
Greenish yellow |
| Tvr24 | Indeterminate |
Semi-erect |
Greenish purple |
Cuneate |
M |
+ |
GN |
Greenish purple |
Medium |
Medium |
1 |
Greenish gray |
Greenish yellow |
| Tvr28 | Indeterminate |
Erect |
Greenish purple |
Cuneate |
M |
+ |
GN |
Greenish purple |
Medium |
Long |
1 |
Greenish gray |
Greenish yellow |
| Tvr32 | Indeterminate |
Erect |
Greenish purple |
Deltoid |
M |
+ |
GN |
Green |
Medium |
Long |
1 |
Greenish gray |
Greenish yellow |
| Tvr49 | Indeterminate |
Semi-erect |
Greenish purple |
Deltoid |
M |
+ |
GN |
Greenish purple |
Medium |
Long |
1 |
Greenish purple |
Greenish yellow |
| Tvr65 | Indeterminate |
Erect |
Greenish purple |
Cuneate |
M |
+ |
GN |
Greenish purple |
Medium |
Short |
1 |
Green |
Greenish yellow |
| Tvr79 | Indeterminate |
Semi-erect |
Purple |
Cuneate |
M |
+ |
GN |
Green |
Long |
Medium |
1 |
Greenish purple |
Greenish purple |
| Tvr83 | Indeterminate |
Spreading |
Green |
Ovate |
S |
+ |
DGN |
Purple |
Short |
Short |
2 |
Greenish yellow |
Bluish yellow |
| Tvr34 | Indeterminate |
Erect |
Green |
Deltoid |
M |
+ |
GN |
Greenish purple |
Short |
Medium |
1 |
Greenish purple |
Greenish purple |
| 1: Above canopy, 2: Below canopy, L: Large, M: Medium, S: Small, GN: Green, DGN: Dark green and +: Present | |||||||||||||
| Table 2b: | Morphological characteristics of ten mungbean (Vigna radiata L.) accessions in derived savanna agro-ecology of Southeastern Nigeria | |||
| Accession | Immature pod color |
Pod shape |
Attachment of pod peduncle |
Pod pubescence |
Pod curvature |
Flower color |
Seed color |
Lustre on seed surface |
Seed shape |
Constriction of pods between seeds |
| Tvr18 | Deep green |
Round |
Intermediate |
Intermediate |
Medium |
Pale yellow |
Mixed |
Dull |
Round |
Present |
| Tvr19 | Light green |
Round |
Intermediate |
Intermediate |
Least curved |
Light yellow |
Mixed |
Dull |
Round |
Present |
| Tvr24 | Light green |
Round |
<90 |
Intermediate |
Least curved |
Pale yellow |
Mixed |
Dull |
Round |
Present |
| Tvr28 | Light green |
Round |
<90 |
Heavily pubescent |
Least curved |
Pale yellow |
Mixed |
Dull |
Round |
Present |
| Tvr32 | Light green |
Round |
Intermediate |
Heavily pubescent |
Least curved |
Pale yellow |
Mixed |
Dull |
Round |
Present |
| Tvr49 | Light green |
Round |
Intermediate |
Heavily pubescent |
Least curved |
Pale yellow |
Mixed |
Dull |
Round |
Present |
| Tvr65 | Light green |
Round |
Intermediate |
Heavily pubescent |
Least curved |
Pale yellow |
Mixed |
Dull |
Round |
Present |
| Tvr79 | Light green |
Round |
Intermediate |
Heavily pubescent |
Least curved |
Pale yellow |
Mixed |
Dull |
Round |
Present |
| Tvr83 | Deep green |
Round |
Intermediate |
Heavily pubescent |
Least curved |
Bright yellow |
Golden brown |
Shiny |
Oval |
Present |
| Tvr34 | Light green |
Round |
Intermediate |
Heavily pubescent |
Least curved |
Pale yellow |
Mixed |
Dull |
Round |
Present |

|
Variation in agronomic traits of ten mungbean accessions evaluated in derived savanna agro-ecology of Southeast Nigeria
Growth: Variations were observed among the ten genotypes studied. Except Tvr83 and Tvr24 which produced shorter plants, averaging peak heights of 30.00 and 36.88 cm, respectively.
At 8 WAP, the other 8 genotypes were statistically similar in height with Tvr65 recording the tallest plant (64.56 cm) (Fig. 1b). The Tvr83 and Tvr24 produced consistently shorter plants across the weeks of investigation. Leaf numbers did not significantly vary at 2 and 4 WAP (Fig. 1c). A significant effect of genotype became evident at 6, 8 and 10 WAP with Tvr83 producing the highest leaf number (65.38) compared to Tvr32 (35.71) which was the least. All genotypes showed comparable NOB at 2 WAP. However, a significant effect was observed at 4, 6, 8 and 10 WAP, with Tvr83 recording higher (p<0.05) NOB (15.08) in contrast to Tvr32 (5.53) which was the least (Fig. 1d). A significant effect of genotype on leaf area was recorded across all weeks of investigation.

|

|
Tvr83 produced consistently thinner and smaller leaves throughout the study with a mean peak of 59.66 cm2 at 10 WAP compared to Tvr49 with the highest mean of 209.27 cm2 (Fig. 1e). Stem diameter was not affected by genotype at 2 WAP. However, at 4, 6, 8 and 10 WAP, the genotypic effect became evident. The Tvr19 and Tvr79 showed consistent and progressive superiority in stem diameter across the weeks of study. The widest stem diameter was recorded in Tvr79 (1.42 cm) while Tvr83 (0.62 cm) was the thinnest at 10 WAP (Fig. 1f).
| Table 3: | Variation in yield and yield components of ten mung bean accessions evaluated in derived savanna agro-ecology of Southeast Nigeria | |||
Geno type |
Number of pods/plant |
Pod length (cm) |
Pod width (cm) |
Number of seeds/pod |
Number of seeds/plant |
Seed length (cm) |
Seed width (cm) |
Seed weight/ plant (g) |
100-seed weight (g) |
Seed yield (t ha–1) |
Tvr18 |
27.27±8.96 |
8.91±0.05 |
0.40±0.02 |
13.5±0.50 |
227.86±38.87 |
0.47±0.01 |
0.38±0.02 |
9.29±0.77 |
5.41±0.60 |
3.10±0.54 |
Tvr19 |
16.33±10.60 |
8.28±0.42 |
0.37±0.04 |
13.25±1.00 |
166.08±139.09 |
0.40±0.01 |
0.35±0.00 |
7.01±6.36 |
5.10±0.51 |
2.34±0.81 |
Tvr24 |
22.75±14.78 |
8.47±0.40 |
0.38±0.03 |
12.58±0.38 |
181.58±83.60 |
0.44±0.00 |
0.35±0.01 |
6.40±3.69 |
4.78±0.58 |
2.13±0.74 |
Tvr28 |
13.42±9.78 |
8.95±0.28 |
0.41±0.08 |
13.5±0.66 |
147.17±112.70 |
0.44±0.01 |
0.35±0.01 |
5.47±4.68 |
4.81±0.62 |
1.82±1.10 |
Tvr32 |
11.28±2.43 |
8.6±0.54 |
0.43±0.05 |
11.69±1.17 |
91.31±21.50 |
0.49±0.03 |
0.40±0.01 |
4.88±2.41 |
4.80±0.80 |
1.63±0.85 |
Tvr49 |
12.00±11.14 |
8.92±0.76 |
0.45±0.04 |
11.44±2.99 |
136.83±129.41 |
0.48±0.03 |
0.40±0.00 |
6.05±5.96 |
5.90±0.35 |
2.02±0.92 |
Tvr65 |
15.58±10.34 |
8.62±0.51 |
0.40±0.04 |
9.75±3.38 |
174.83±190.70 |
0.49±0.02 |
0.39±0.03 |
7.43±8.20 |
5.50±0.20 |
2.48±1.40 |
Tvr79 |
26.58±15.60 |
8.75±0.75 |
0.44±0.01 |
12.58±1.38 |
213.92±118.71 |
0.48±0.02 |
0.39±0.01 |
8.94±5.61 |
5.35±0.41 |
2.98±0.55 |
Tvr83 |
32.61±7.50 |
4.75±0.06 |
0.36±0.04 |
7.06±0.42 |
164.33±45.52 |
0.49±0.01 |
0.37±0.02 |
6.60±2.10 |
5.37±0.27 |
2.20±1.54 |
Tvr92 |
9.33±1.89 |
6.87±0.52 |
0.45±0.10 |
7.50±2.12 |
69.67±23.57 |
0.59±0.00 |
0.41±0.03 |
3.03±0.10 |
4.98±0.55 |
1.01±1.02 |
LSD0.05 |
14.76 |
0.83 |
NS |
2.98 |
NS |
0.06 |
0.02 |
NS |
NS |
0.26 |
Mean |
18.72 |
8.11 |
0.41 |
11.29 |
157.36 |
0.47 |
0.38 |
6.51 |
5.2 |
2.17 |
SD |
8 |
1.33 |
0.03 |
2.4 |
49.17 |
0.03 |
0.02 |
1.85 |
0.37 |
0.62 |
CV% |
42.77 |
16.38 |
8.02 |
21.25 |
31.25 |
7 |
6.02 |
28.42 |
7.09 |
28.42 |
| NS: Not significantly different at p<0.05 according LSD mean separation procedure, LSD: Least significant difference between two means, SD: Standard deviation and CV%: Percent coefficient of variation | ||||||||||
|
Yield: Variation in yield and yield components were recorded among the ten accessions (Table 3). Of the 10 yield traits evaluated, the genotypes varied significantly (p<0.05) in 6 traits (NOP per plant, pod length plant per plant, NOS per pod, seed length, seed width and seed yield) and were comparable in 4 traits (p>0.05). The NOP per plant ranged from 9.33 in Tvr92 to 27.27 in Tvr18. Pod length ranged from 4.75 in Tvr83 to 8.95 in Tvr28. Pod width ranged from 0.36 cm in Tvr83 to 0.45 cm in Tvr49. The NOS per pod ranged from 7.06 in Tvr83 to 13.5 in Tvr18. NOS per plant ranged from 69.67 in Tvr92 to 227.86 in Tvr18.
Seed length ranged from 0.40 cm in Tvr19 to 0.59 cm in Tvr92. Seed width ranged from 0.35 cm in Tvr24 to 0.41 cm in Tvr92. Seed weight per plant ranged from 3.03 g in Tvr92 to 9.29 in Tvr18. Weight of 100-seed ranged from 4.78 g in Tvr24 to 5.90 g in Tvr49. Seed yield ranged from 1.01 t ha–1 in Tvr92 to 3.10 t ha–1 in Tvr18. Coefficient of variation ranged from 6.03 for seed width to 42.77% for NOP per plant.
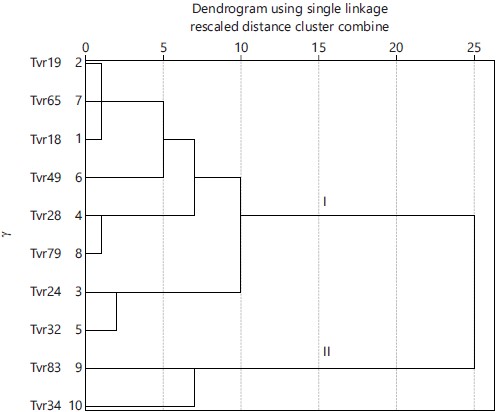
Cluster analysis using single linkage method grouped the ten accessions into 2 clusters at a Euclidian distance >10. Accessions Tvr19, Tvr65, Tvr18, Tvr49, Tvr28, Tvr79, Tvr24 and Tvr32 occupied cluster I while, Tvr34 and Tvr83 occupied cluster II (Fig. 2).
DISCUSSION
The accessions differed significantly in terms of morphological and agronomic characteristics. Variations in morphological attributes such as growth pattern, hypocotyl colour, leaf shape and colour, petiole colour, raceme position, point of attachment of peduncle, pod pubescence, etc. (Table 2a and b) could suggest differences in their DNA sequences and karyotypes21, an indication that the accessions may have evolved from ecologically diverse habitats6, ancestry or at different evolutionary stages of development. Variations in phenological attributes including NOD to 1st and 50% emergence and NOD to 1st and 50% flowering, NOD to 1st and 50% podding are very important parameters that could serve as morphological markers to discriminate against late maturing genotypes. The existence of genetic variability is the basis upon which crop improvement is advanced22. It is pivotal to the success of any breeding program23, as the availability of sufficient variability in a germplasm collection can almost nearly guarantee success24. Three accessions, Tvr32, Tvr49 and Tvr83 showed consistency in earliness in terms of NOD to emergence, flowering and podding and are therefore early maturing genotypes while two accessions, Tvr18 and Tvr65 were consistently late and are classified as late maturing. The other accessions, Tvr19, Tvr24, Tvr28, Tvr79 and Tvr34 were neither early nor late maturing.
Variation in plant height, leaf number, leaf area, pod and seed traits among the ten mungbean accessions suggests differences in their genetic makeup. Accessions Tvr18 and Tvr79 were clustered together in cluster I and were the best in yield traits inferring a common ancestry. The coefficient of variation revealed that NOP plant (42.77%), NOS pod (31.25%) and seed yield per hectare (28.42%) were the most discriminatory yield parameters. Based on CV%, Tvr18 and Tvr79 which recorded higher numbers of seeds per plant, NOS pod and seed yield per hectare were the best-yielding accessions. These were closely followed by Tvr65, Tvr19 and Tvr83, respectively. However, Tvr83 was unique from the other genotypes as it produced distinctively the smallest leaf area, shiny seeds, higher leaf numbers and higher NOP plant (p<0.05) relative to the rest genotypes which suggests that it could be a distant relative of the other accessions as elucidated by the dendrogram diagram (Fig. 2) thus, making it an excellent candidate for mungbean hybridization and introgression programs, especially in traits for which it showed superiority.
Clustering of genotypes helps in partitioning a large heterogeneous set of genotypes into smaller and relatively more homogeneous clusters25. It is important in identifying distant relatives of a species which is a requirement for crop improvement through hybridization22,26 as plant breeders are presented with ample opportunities to create new, improved cultivars through genetic variability27. Closely related genotypes must have clustered together in clusters I and II. This is consistent with28,29.
Implications Tvr18 and Tvr79 which were the best yielding genotypes could potentially provide flatulence-free proteins to the Nigerian populace while Tvr83 is a valuable candidate for improvement of the branching potentials of Tvr18 and Tvr79 through hybridization. However, further evaluation in multilocation trials for yield stability needs to be carried out.
CONCLUSION
The study characterized and evaluated 10 mungbean accessions based on 23 morphological and 16 agronomic parameters in the derived savanna agroecological zone of Southeast Nigeria. Morphological and agronomic variations were recorded for most of the traits investigated. Two accessions, Tvr18 and Tvr79 performed better in terms of NOP per plant, NOS per pod and seed yield which was the three most varied yield parameters and were therefore selected for advancement to the next generation. The ten accessions were clustered into two distinct groups, cluster I and II, with 8 and 2 accessions, respectively, suggesting that accessions clustered together are close relatives. The Tvr83 which was grouped with Tvr34 in cluster II recorded the highest NOL, NOB and NOP per plant in addition to having smaller leaves compared to the other nine accessions. Hence, it could serve as a promising donor for the improvement of Tvr18 and Tvr79 in any of these traits for which it showed superiority.
SIGNIFICANCE STATEMENT
The purpose of this study was to identify high yielding mungbean accessions to add to the legume base of southeast Nigeria in an attempt to secure cheap alternative plant protein sources and in cushioning the effect of hunger and malnutrition in the region in line with the UN Sustainable Development Goals. Two high yielding accessions, Tvr18 and Tvr79 and one high branching type were identified. These high yielding accessions could potentially provide flatulence free proteins to the Nigerian populace while the high branching type is a valuable candidate for improvement of the branching potentials of the other two through hybridization.
ACKNOWLEDGMENT
The authors acknowledge the Genetic Resource Center, International Institute of Tropical Agriculture for making the seeds available.
REFERENCES
- Chukwudi, U.P., S. Mavengahama, F.R. Kutu and L.E. Motsei, 2022. Heat stress, varietal difference, and soil amendment influence on maize grain mineral concentrations. Agriculture, 12: 1633.
- van Dijk, M., T. Morley, M.L. Rau and Y. Saghai, 2021. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010-2050. Nat. Food, 2: 494-501.
- Agugo, B.A.C., P.C. Oguike and B.O. Kanu, 2010. A preliminary field assessment of mungbean (Vigna radiata L. Wilczek) yield in the rain forest zone of Southeastern Nigeria. Am. Eurasian J. Agric. Environ. Sci., 8: 752-757.
- Kumar, J., A.K. Choudhary, R.K. Solanki and A. Pratap, 2011. Towards marker-assisted selection in pulses: A review. Plant Breed., 130: 297-313.
- Evgenidis, G., E. Traka-Mavrona and M. Koutsika-Sotiriou, 2011. Principal component and cluster analysis as a tool in the assessment of tomato hybrids and cultivars. Int. J. Agron., 2011: 697879.
- Sangiri, C., A. Kaga, N. Tomooka, D. Vaughan and P. Srinives, 2007. Genetic diversity of the mungbean (Vigna radiata, Leguminosae) genepool on the basis of microsatellite analysis. Aust. J. Bot., 55: 837-847.
- Ramakrishnan, M.N., R. Schafleitner and S.H. Lee, 2020. The Mungbean Genome. 1st Edn., Springer, Cham, Switzerland, ISBN: 978-3-030-20007-7, Pages: 191.
- Ngwuta, A.A., A.C. Okoro, B.A.C. Agugo, A.C. Odiyi, K.A. Eke and I.J. Ogoke, 2010. Variations in some morphological and yield characteristics of mungbean (Vigna radiata (L.) Wilczek) cultivars introduced into the rain-forest agroecology of Southeastern Nigeria. Int. J. Agric. Rural Dev., 13.
- Okweche, S.I. and T.R. Avav, 2013. Yield evaluation of some cultivars of mungbean (Vigna radiata (L) Wilczek) in Southern Giunea Savanna location of Nigeria. Int. J. Plant Anim. Environ. Sci., 3: 85-88.
- Akintoye, H.A., M.T. Olatunji, A.O. Shokalu, A.G. Adebayo and E.M. Okoyo, 2022. Triggering a mungbean transformation in East and West Africa: Evaluation of adaptability of fifteen lines of mungbean in Southwestern Nigeria. Acta Hortic., 1348: 277-284.
- Ihejiofor, P.N., U.N. Ukwu and G. Adeoye, 2022. Determination of Kolgrace bio-fertilizer rate for optimum greengram (Vigna radiata L. Wilczek) production in Ibadan, Southwest Nigeria. Agro-Science, 21: 82-87.
- Ihejiofor, P.N., U.N. Ukwu and G.O. Adeoye, 2020. Comparative effects of different levels of kolgrace organic fertilizer on the growth and yield attributes of greengram (Vigna radiata (L) Wilczek) in the screenhouse. Asian J. Res. Agric. For., 6: 1-7.
- Dahiya, P.K., M.J.R. Nout, M.A. van Boekel, N. Khetarpaul, R.B. Grewal and A. Linnemann, 2014. Nutritional characteristics of mung bean foods. Br. Food J., 116: 1031-1046.
- Arumugam, R., S. Rajasekaran and S.M. Nagarajan, 2010. Response of arbuscular mycorrhizal fungi and Rhizobium inoculation on growth and chlorophyll content of Vigna unguiculata (L) Walp Var. Pusa 151. J. Appl. Sci. Environ. Manage., 14: 113-115.
- Tang, D., Y. Dong, H. Ren, L. Li and C. He, 2014. A review of phytochemistry, metabolite changes, and medicinal uses of the common food mung bean and its sprouts (Vigna radiata). Chem. Cent. J., 8.
- Nadeem, M.A., R. Ahmad and M.S. Ahmad, 2004. Effect of seed inoculation and different fertilizer levels on the growth and yield of mungbean (Vigna radiata L.). J. Agron., 3: 40-42.
- Mandal, S., M. Mandal, A. Das, B. Pati and A. Ghosh, 2009. Stimulation of indoleacetic acid production in a Rhizobium isolate of Vigna mungo by root nodule phenolic acids. Arch. Microbiol., 191: 389-393.
- Kim, D.K., S.C. Jeong, S. Gorinstein and S.U. Chon, 2012. Total polyphenols, antioxidant and antiproliferative activities of different extracts in mungbean seeds and sprouts. Plant Foods Hum. Nutr., 67: 71-75.
- Kaur, R., A.K. Toor, G. Bassi and T.S. Bains, 2017. Characterization of mungbean (Vigna radiata L. Wilczek) varieties using morphological and molecular descriptors. Int. J. Curr. Microbiol. Appl. Sci., 6: 1609-1618.
- Hamid, A. and W. Agata, 1989. Estimating leaf area in mungbean (Vigna radiata). J. Agric. Sci., 113: 165-167.
- Agbo, C.U. and N.U. Ukwu, 2010. Morphology and chromosome numbers of Gongronema latifolia benth. clones from Nigeria. Afr. Crop Sci. J., 18: 29-38.
- Gokulakrishnan, J., B.S. Kumar and M. Prakash, 2023. Studies on genetic diversity in mung bean (Vigna radiata L.). Legume Res., 35: 50-52.
- Ukwu, N.U. and B. Olasanmi, 2018. Crossability among five cassava (Manihot esculenta CRANTZ) varieties. Mod. Concepts Dev. Agron., 2: 183-188.
- Ebert, A.W. and J.M.M. Engels, 2020. Plant biodiversity and genetic resources matter! Plants, 9: 1706.
- Rana, J.C., T.R. Sharma, R.K. Tyagi, R.K. Chahota and N.K. Gautam et al., 2015. Characterisation of 4274 accessions of common bean (Phaseolus vulgaris L.) germplasm conserved in the Indian gene bank for phenological, morphological and agricultural traits. Euphytica, 205: 441-457.
- John, K.N.B., D. Packiaraj, M. Pandiyan and N. Senthil, 2015. Tailoring genetic diversity of mungbean (Vigna radiata (L.) Wilczek) germplasm through cluster analysis for yield and yield related traits. Trends Biosci., 8: 3239-3244.
- Bhandari, H.R., A.N. Bhanu, K. Srivastava, M.N. Singh and H.A. Shreya, 2017. Assessment of genetic diversity in crop plants-An overview. Adv. Plants Agric. Res., 7: 279-286.
- Gayacharan, K. Tripathi, S.K. Meena, B.S. Panwar, H. Lal, J.C. Rana and K. Singh, 2020. Understanding genetic variability in the mungbean (Vigna radiata L.) genepool. Ann. Appl. Biol., 177: 346-357.
- Mwangi, J.W., O.R. Okoth, M.P. Kariuki and N.M. Piero, 2021. Genetic and phenotypic diversity of selected Kenyan mung bean (Vigna radiata L. Wilckzek) genotypes. J. Genet. Eng. Biotechnol., 19: 142.
How to Cite this paper?
APA-7 Style
Ukwu,
U.N., Oburu,
B.N., Muojiama,
S.O., Osadebe,
V.O., Dauda,
N., Adewuyi,
S.O. (2023). Morphology and Agronomic Evaluation of Ten Mungbean (Vigna radiata L.) Accessions in Derived Savanna Agroecology of Southeast Nigeria. Asian Journal of Biological Sciences, 16(2), 110-120. https://doi.org/10.3923/ajbs.2023.110.120
ACS Style
Ukwu,
U.N.; Oburu,
B.N.; Muojiama,
S.O.; Osadebe,
V.O.; Dauda,
N.; Adewuyi,
S.O. Morphology and Agronomic Evaluation of Ten Mungbean (Vigna radiata L.) Accessions in Derived Savanna Agroecology of Southeast Nigeria. Asian J. Biol. Sci 2023, 16, 110-120. https://doi.org/10.3923/ajbs.2023.110.120
AMA Style
Ukwu
UN, Oburu
BN, Muojiama
SO, Osadebe
VO, Dauda
N, Adewuyi
SO. Morphology and Agronomic Evaluation of Ten Mungbean (Vigna radiata L.) Accessions in Derived Savanna Agroecology of Southeast Nigeria. Asian Journal of Biological Sciences. 2023; 16(2): 110-120. https://doi.org/10.3923/ajbs.2023.110.120
Chicago/Turabian Style
Ukwu, Uchenna, Noble, Blessing Ngozika Oburu, Stella Ogochukwu Muojiama, Vivian Ogechi Osadebe, Nathaniel Dauda, and Solomon Oluwaseyi Adewuyi.
2023. "Morphology and Agronomic Evaluation of Ten Mungbean (Vigna radiata L.) Accessions in Derived Savanna Agroecology of Southeast Nigeria" Asian Journal of Biological Sciences 16, no. 2: 110-120. https://doi.org/10.3923/ajbs.2023.110.120

This work is licensed under a Creative Commons Attribution 4.0 International License.