Immune Responses Elicited by COVID-19 Vaccines
-
Kaushik Bharati


COVID-19 Expert Committee-UNESCO, New Delhi, Delhi 110021, India
| Received 09 Sep, 2022 |
Accepted 15 Dec, 2022 |
Published 31 Mar, 2023 |
Coronavirus disease 2019 (COVID-19) vaccines have been developed in record time-the fastest in medical history. Many types of vaccines, using novel platforms, have been developed. Many of these vaccines are already being used in vaccination drives worldwide. Many more are still in the pipeline to strengthen and replenish the vaccine armamentarium. Many immune mechanisms are involved in response to vaccination. The currently available vaccines generate robust humoral and cell-mediated immune responses and exhibit high levels of safety and efficacy. The neutralizing antibodies (NAbs) in particular, play a crucial role in blocking viral transmission. This review explored the various types of immune responses elicited by different types of COVID-19 vaccines and their implications in thwarting the spread of the virus.
| Copyright © 2023 Kaushik Bharati. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Coronavirus disease 2019 (COVID-19) is so-called because it was first reported on 2019-31st December, 2019 to be more precise. On this day, a cluster of atypical pneumonia cases of unknown etiology was reported from the city of Wuhan in Hubei Province, China to the World Health Organization (WHO). The following day i.e., January 1st, 2020 (New Year’s Day), the WHO swung into action by activating its emergency response team. This was to be the beginning of a long-haul, harrowing experience for the entire world, which still shows no signs of abating. The WHO anticipated that the threat of a pandemic was very real. The WHO Director-General, Dr. Tedros Adhanom Ghebreyesus warned about it as early as 2017. The UN organization even drew up a blueprint to prioritize research in the area of emerging diseases. This research blueprint labelled a hitherto unidentified emerging disease with pandemic potential as “Disease X”. Now, COVID-19 has emerged to be “Disease X”. A summary of the timeline of WHO’s response to the pandemic was highlighted in Table 1.
The COVID-19 is caused by the Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), which belongs to the family Coronaviridae and sub-family Betacoronavirus. This virus has been known to infect mammals, including mice, bats and pangolins. But this is the first time it has infected humans and till August 27th, 2021, it was responsible for 215 million cases and 4.48 million deaths across 221 countries and territories worldwide1.
|
Since the beginning of the pandemic, it was relatively clear in the minds of the scientists that a vaccine would be the answer to taming the disease. However, it was not clear what type of immune responses would be generated and what the immune correlates of protection would be. The COVID-19 vaccine development heavily relied on the knowledge gained from the previous SARS epidemic of 2002-2003 and the Middle East respiratory syndrome (MERS) epidemic of 2012. Currently approved COVID-19 vaccines are based on the experience with SARS-CoV (renamed as SARS-CoV-1 by WHO to distinguish it from the current SARS-CoV-2. This abbreviation will be followed in this review) and Middle East respiratory syndrome coronavirus (MERS-CoV). However, it should be noted that although candidate vaccines against these pathogens had been developed, they didn’t go beyond Phase 1 clinical trials2.
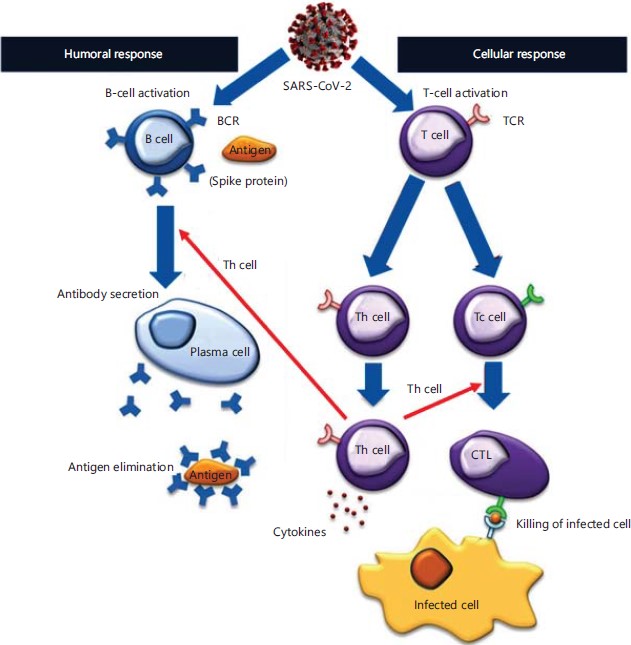
Types of immunity relevant to vaccines3: In the context of vaccines, both arms of the adaptive immune response, namely humoral and cellular immunity, are essential. This is true for all viral vaccines4 (Fig. 1).
The active components of the humoral immune response are antibodies, which are generated by immunization with an antigen that is specific to the pathogen in question. In the case of COVID-19, the specific antigen is the spike (S) protein of SARS-CoV-2. Consequently, the generated antibodies bind to the virus, thereby preventing it from entering the cell and further transmitting the infection.

|
| Table 1: | Timeline of events and WHO’s response to the pandemic | |||
| Date | WHO’s response |
| December 31st, 2019 | Cluster of atypical pneumonia cases of unknown etiology reported from Wuhan, China |
| January 1st, 2020 | WHO activated its Incident Management Support Team |
| January 2nd, 2020 | WHO alerted the Global Outbreak Alert and Response Network (GOARN) |
| January 11th, 2020 | First genomic sequence of SARS-CoV-2 shared by China |
| First death from COVID-19 reported from China | |
| January 13th, 2020 | First COVID-19 case reported outside China (Thailand) |
| January 22nd, 2020 | WHO confirmed human-to-human transmission of SARS-CoV-2 |
| January 30th, 2020 | WHO declared COVID-19 outbreak as a Public Health Emergency of International Concern (PHEIC) |
| February 11th, 2020 | 2019 novel coronavirus (2019-nCoV) renamed as COVID-19 |
| March 11th, 2020 | WHO declared COVID-19 as a ‘pandemic’ |
The active components of cellular immunity are the T-cells, so-called because they are processed in the thymus gland. Like humoral immunity, T-cells are also activated by immunization. There are two major subsets of T-cells, namely, CD4+ and CD8+ T-cells. The majority of T-cells are of the former type. Their primary function is to recognize antigens presented on the cell surface, usually in the context of major histocompatibility complex (MHC) molecules. Hence, as opposed to B-cells, which recognize soluble antigens, T-cells can only recognize antigens displayed on the surface of host cells. There are various subsets of T-cells, all of which work in a concerted manner to generate a robust cellular immune response. The memory T-cells are especially important in the context of vaccines, as they retain a memory of the antigen (in this case SARS-CoV-2) so that if exposed to the virus again, it immediately alerts the other T-cells that generate an aggressive cellular immune response, which effectively neutralizes the virus, thereby preventing onward transmission.
SARS-CoV-2 spike protein
Target antigen for all candidate vaccines: The aetiologic agent of COVID-19, namely, SARS-CoV-2 is a single-stranded, positive-sense RNA virus, having a genome size of approximately 29.9 kb5. This was in contrast to SARS-CoV-1 and MERS-CoV which have genome sizes of approximately 27.9 kb and 30.1 kb, respectively6,7. The SARS-CoV-2 has four structural proteins, namely, spike (S), envelope (E), membrane (M) and nucleocapsid (N). While the first three are present on the surface, the fourth one is present inside, bound to the RNA, thereby protecting it from degradation8. The S protein plays a pivotal role in eliciting the immune response. It is a trimeric class I fusion protein that binds to the Angiotensin-Converting Enzyme 2 (ACE2) receptor on the cell surface, which facilitates the fusion of the virus and subsequent entry into the cell. The S protein has two subunits-S1 and S2 that mediate receptor binding and fusion, respectively. The S1 subunit contains the receptor-binding domain (RBD), which binds to the ACE2 receptor9. Hence, the primary focus of scientists was to develop a vaccine that elicits an immune response targeted specifically against the RBD, which would confer sterilizing immunity, thereby completely preventing infection (Fig. 2).

|
Immunodominant epitopes
Key to optimization of vaccine design: Optimally designed candidate vaccines having immunodominant epitopes are of paramount importance for generating a robust immune response. Moreover, excluding unnecessary and non-immunogenic domains will ensure that harmful side effects are minimized. Hence, the search for immunodominant B- and T-cell epitopes was a rigorous exercise in the initial stages of vaccine design.
As whole-genome sequences started emerging, it was seen that SARS-CoV-2 shared around 79% similarity with SARS-CoV-1, which simplified the vaccine design process. This high level of similarity between the two viruses led scientists to predict several immunogenic B- and T-cell epitopes on the surface of SARS-CoV-210. The 49 linear and 6 discontinuous B-cell epitopes, as well as several T-cell epitopes were discovered, most of which were from the S protein11. Additionally, bioinformatics approaches led to the discovery of 25 immunogenic epitopes from SARS-CoV-2, that covered almost 87% of the global population. Moreover, these were safe as they didn’t elicit any allergic reaction or exhibit toxicity and had a negligible chance for the development of autoimmunity. Hence, these were chosen as possible candidates for designing vaccines12.
Types of COVID-19 vaccine platforms: As of August 28th, 2021 there were 296 candidate vaccines under development, including 112 in clinical trials and 184 in pre-clinical development in various animal models. These include a whole gamut of vaccine platforms, encompassing traditional platforms (killed/inactivated and live-attenuated vaccines), recombinant protein platforms (protein subunit and VLP vaccines), viral vector platforms (replicating and non-replicating), as well as more advanced ones, such as nucleic acid platforms (DNA and RNA), to name a few in Fig. 313. Many of these candidate vaccines are already approved and are currently being used worldwide for mass vaccination. Several of these vaccines have received WHO’s Emergency Use Listing (EUL), which is a stamp of quality, concerning vaccine safety and efficacy. As of August 19th, 2021, these EUL-approved vaccines included Pfizer/BioNTech and Moderna (mRNA platforms), Oxford University/AstraZeneca and Janssen (non-replicating viral vector platforms), Sinopharm and Sinovac (inactivated vaccine platforms). Besides these, there are currently thirteen other candidate vaccines under evaluation for WHO approval14.
These vaccines were developed in a record-breaking time-the fastest in medical history. Needless to say, the major credit goes to the scientists, doctors and vaccine manufacturers for their tireless efforts. Having said this, much of the groundwork was laid by the Coalition for Epidemic Preparedness Innovations (CEPI), a global partnership, which was launched in 2017 with the sole purpose of developing the various vaccine platforms, so that they would be ready for use in the event of the emergence of a deadly pathogen, such as the current SARS-CoV-2. Hence, the development of these vaccines is truly due to global collaborative efforts, the magnitude and intensity of which have never been seen before in the entire history of scientific research.

|
Immune responses induced by vaccination against SARS-CoV-215: As discussed above, vaccines generate two types of the immune responses, namely, humoral and cell-mediated. Humoral immunity is mediated by antibodies, which are again of two types-binding and neutralizing antibodies (NAbs). The current vaccines are largely being evaluated for their antibody-inductive capacity. Most of these are binding antibodies, meaning that they simply bind to the S protein. On the contrary, the NAbs bind to the RBD of the S protein and neutralize the viral activity. Therefore, the latter class of antibodies is much more useful in assessing vaccine efficacy and are usually detected using a plaque reduction neutralization test with a 50% cut-off (PRNT50).
Antibodies have traditionally been considered the gold standard for ascertaining correlates of protection, as opposed to cell-mediated immune responses. The main reason is the ease and cost-effectiveness of the procedures used. Measurement of cell-mediated immunity is time-consuming, cumbersome and requires expensive instruments. It is often measured using intracellular cytokine staining (ICS) assay or enzyme-linked immunospot (ELISpot) assay. The latter uses the S protein to stimulate the peripheral blood mononuclear cells (PBMCs), which is a prerequisite for conducting this assay. Despite the barriers posed by these high-end procedures, they yield highly useful data that buttress and expand the scope of the data generated by the antibody assays. The presence of memory cells is particularly useful for predicting vaccine efficacy upon future exposure to the virus.
Major candidate vaccines under different stages of development: There are a total of 296 candidate vaccines under different stages of development, of which 112 are undergoing clinical trials. Of the various types of vaccines, the protein subunit vaccine is the front-runner, with a total of 38 candidates. Others include mRNA (18 candidates), non-replicating viral vector (17 candidates), inactivated virus (16 candidates) and DNA (11 candidates), among several others. The top 8 front-runners were presented in Fig. 4.
Protein subunit vaccines: As the name suggests, this type of vaccine utilizes a protein subunit of the virus16, which in this case is the S protein. This vaccine platform has, by far, the largest number of vaccine candidates in clinical trials and pre-clinical development. The most advanced protein subunit vaccine i.e., the one developed by Novavax was discussed below.
Novavax vaccine: This vaccine was developed by Novavax, a company based in Gaithersburg, Maryland, USA. It is a two-dose vaccine that is administered 21 days apart and is technically termed NVX-CoV2373. The recombinant S protein contains mutations at the S1/S2 cleavage sites that confer protease resistance and is formulated with the so-called adjuvant. This adjuvant consists of Quillaja saponins formulated with cholesterol and phospholipids into nanoparticles, which are known to elicit both Th1 and Th2 immune responses, as well as induce antibodies of different subclasses. With this novel adjuvant, the anti-S IgG titers were 63,160 enzyme-linked immunosorbent assay (ELISA) units and NAb titers were as high as 3,906. When tested without the adjuvant, the vaccine-elicited much lower levels of antibodies17.
The vaccine generated a CD4+ T-cell response, which became evident 7 days after the second dose. The cytokine milieu indicated that the cellular immune response was more skewed towards Th1 (IFN-γ, IL-2 and TNF), rather than Th2 (IL-5 and IL-13)17.
The Novavax vaccine was recently evaluated in a Phase 3 randomized, observer-blinded, placebo-controlled trial conducted among 14,039 individuals in the UK. The vaccine was safe and exhibited an efficacy of 86.3% against the alpha variant (B.1.1.7) and 96.4% against non-alpha variants (beta, gamma and delta variants). Reactogenicity was mild and transient and the incidence of severe adverse events following immunization (AEFI) was low18.
mRNA vaccines: The mRNA vaccines are based on messenger RNA technology and along with the DNA vaccines, are collectively known as nucleic acid or genetic vaccines. The two leading mRNA vaccines that have received WHO’s EUL approval and are currently being used across the globe, namely the Pfizer/BioNTech and Moderna vaccines, are discussed below.
Pfizer/BioNTech vaccine: This is a two-dose mRNA vaccine that is delivered 21 days apart. Pfizer/BioNTech initially developed two vaccine constructs, which they termed BNT162b1 and BNT162b2. Both of them were similar, except that the former encoded the secreted trimerized RBD of the S protein, while the latter encoded the membrane-anchored full-length S protein, stabilized in the prefusion conformation. These two constructs were tested in a randomized, placebo-controlled, observer-blinded, dose-escalation, Phase 1 trial in the US. There were two age groups: 18 to 55 years and 65 to 85 years, who received either a placebo or any one of the two, lipid nanoparticle (LNP) formulated mRNA constructs. In the case of BNT162b2, there was a much lower incidence and severity of AEFI compared to BNT162b1. However, both vaccine constructs elicited similar levels of NAb titers in younger and older adults. These titers were higher than those of convalescent serum samples. Due to the milder side effects of BNT162b2, this candidate vaccine was taken forward for further clinical development. Notably, this was the first published clinical trial of an mRNA vaccine19. In a subsequent Phase 3 clinical trial with over 44,000 participants, BNT162b2 showed a staggering efficacy of 91.3% against COVID-19, even after 6 months following the second dose20.
Concerning cell-mediated immunity, strong CD4+ and CD8+ T-cell responses were elicited, which were biased towards the Th1 phenotype. Notably, RBD-specific CD4+ and CD8+ T-cells were significantly elevated. Moreover, IFN-γ and IL-2 secretion were also significantly higher, compared to IL-4, indicating a Th1 polarization21.
Moderna vaccine: This vaccine, like the one by Pfizer/BioNTech, is also an LNP-encapsulated mRNA vaccine, technically termed mRNA-1273. It is a two-dose vaccine that is administered 28 days apart. The mRNA construct encodes the full-length S protein, stabilized in its prefusion conformation. A large-scale Phase 3 randomized, observer-blinded, placebo-controlled clinical trial was conducted at 99 centers across the US, involving 30,420 individuals. The vaccine exhibited an efficacy of 94.1% after the second dose, which was uniform across all groups, including individuals with evidence of SARS-CoV-2 infection and in individuals 65 years of age or older. In the vaccine group, reactogenicity was moderate and transient and serious AEFI was rare. Thus, the vaccine was capable of preventing COVID-19 illness and severe disease22. Humoral immune responses were studied in a Phase 1 dose-escalation, open-label trial including 45 healthy adults 18 to 55 years of age. Two shots were administered 28 days apart. The participants were divided into 3 groups (n = 15 in each group) and three graded doses (25 μg, 100 μg and 250 μg) were administered. The anti-S protein antibodies were higher with increasing doses. The geometric mean titer (GMT) of the antibodies generated in the three groups were 40,227 (25 μg group), 109,209 (100 μg group) and 213,526 (250 μg group), respectively23. The NAb titers also increased significantly following the second dose and were still detectable even after 119 days post-vaccination24.
Cell-mediated immunity was estimated by ICS assay. It was seen that the 25 μg and 100 μg groups elicited CD4+ T-cell responses, which exhibited Th1 polarity as was evident from the cytokine milieu (TNF-α > IL-2 > IFN-γ). However, Th2 cytokine expression (IL-4 and IL-13) was minimal. Moreover, CD8+ T-cell responses were low in the 100 μg group after the second dose23.
DNA vaccines: The DNA vaccines, like mRNA vaccines, are an emerging genetic vaccine technology25 that is being used for several viral diseases26, the most recent being COVID-19. As opposed to mRNA vaccines, which use a single-stranded mRNA molecule, DNA vaccines use a double-stranded DNA molecule. Currently, there are 11 candidate DNA vaccines in clinical trials, of which one has completed all three phases of the trials and has recently been approved for use in India. This DNA vaccine, termed ZyCoV-D, developed by the Indian Company Zydus Cadila, is the world’s first approved DNA vaccine.
DNA vaccine technology, which was first developed in the 1990s, uses DNA encoding the S protein cloned into mammalian expression plasmids. This is a highly promising technology as this vaccine elicits both humoral and cellular immune responses. Moreover, it has several other advantages, such as ease of production, stability at ambient temperatures and convenience in storage and transportation27.
ZyCoV-D
World’s first DNA vaccine28: This vaccine has been developed by Zydus Cadila from India and is the first DNA vaccine in the world to receive emergency use authorization from the Drugs Controller General of India (DCGI) on August 20th, 2021. Although the majority of the vaccines require two doses, ZyCoV-D is a three-dose vaccine, which could be a problem from a logistical standpoint during vaccine deployment and delivery. The vaccine is administered intradermally through a needle-free device in the form of a jet of fluid. The vaccine formulation penetrates the outer epidermal layer of the skin and gets taken up by the dendritic cells in the middle and inner layers, resulting in the expression of the transgene encoded in the plasmid (S protein gene), thereby stimulating the immune system to produce antibodies. This needle-free delivery system, known as PharmaJet Tropis®, has been developed by PharmaJet, which is a company based in Golden, Colorado, USA. This method of delivery is painless as opposed to the standard needle-syringe injection.
The ZyCoV-D has been successfully evaluated in almost 30,000 individuals in all three phases of clinical trials. Moreover, it is the first Indian vaccine to be tested in 12 to 18 years old children and adolescents. The vaccine was found to be well-tolerated, as well as safe and highly immunogenic, both in the adolescent group, as well as the adult group. Following the administration of the third dose, there were no moderate cases in the vaccine group, indicating that the vaccine confers 100% protection against moderate COVID-19 illness. The vaccine exhibited an overall efficacy of 66.6% in a real-time quantitative reverse-transcription polymerase chain reaction (real-time qRT-PCR)-positive COVID-19 cases during an interim analysis.
Inovio vaccine29: A candidate DNA vaccine, termed INO-4800, has been developed by Inovio Pharmaceuticals (Plymouth Meeting, PA, USA) that utilizes the plasmids pGX9501 and pGX9503. These mammalian expression plasmid constructs encode the S protein gene, which is strongly expressed. The INO-4800 generated both cellular and humoral immune responses in mice and guinea pigs just after a single shot. Following the pre-clinical studies, its safety, tolerability and immunogenicity were successfully evaluated in Phase 1 clinical trials and are currently being further evaluated in Phase 2/3 trials. The vaccine is delivered intradermally with the help of electroporation. This technique uses an electrical current to create pores in the cell membrane, which facilitates the entry of the DNA molecules within the cell. The INO-4800 has been shown to induce NAbs that blocked the interaction between the S protein and the ACE2 receptor, thereby halting transmission.
Viral vector vaccines: Viral vector vaccines use an innocuous virus, such as adenovirus to piggyback the transgene encoding the S protein and deliver it into the cell. This concept may be likened to a Trojan horse. There are currently 19 candidate viral vector vaccines in different stages of clinical trials. Seventeen of these are non-replicating viral vector vaccines, while two are replicating viral vector vaccines. Adenovirus-based vaccines are capable of eliciting NAbs, as well as robust cell-mediated immune responses, even after a single dose. It appears that concerning safety and immunogenicity, these vaccines are similar albeit slightly inferior to mRNA and protein subunit vaccines. The major viral vector vaccines are highlighted below.
Oxford University/AstraZeneca vaccine: The major part of the research on this vaccine was done at the University of Oxford and later taken forward by the pharma giant AstraZeneca, headquartered in Cambridge, UK. This vaccine is based on an adenovirus derived from chimpanzees. Hence, this vaccine is technically termed ChAdOx1nCoV-19 (Ch: Chimpanzee, Ad: Adenovirus, Ox: Oxford University and nCoV-19: Novel coronavirus 2019). This adenovirus is non-replicating and thus doesn’t interfere with the expression of the transgene, encoding the S protein along with the tissue plasminogen activator (tPA) leader sequence. An advantage of this chimp adenovirus (ChAd) is that, unlike Ad5, there are negligible levels of ChAd antibodies in humans, as a result of which there is a very little risk posed by pre-existing immunity against this Ad vector15.
A Phase 1/2, single-blind, randomized controlled trial at five trial sites in the UK, showed that the vaccine was safe and antibody titers were significantly boosted after the second dose. Significant T-cell responses were also induced, as measured by IFN-γ ELISpot assays30. Subsequently, three single-blind, randomized clinical trials: (i) Phase 1/2 in the UK, (ii) Phase 2/3 in the UK and (iii) Phase 3 in Brazil and one double-blind Phase 1/2 trial in South Africa were conducted on individuals 18 years and older. The interim analysis of these trials reaffirmed the high degree of safety and efficacy of the vaccine against symptomatic disease. In the group that received two standard doses of the vaccine, the efficacy was 62.1%. However, in the group that inadvertently received a low first dose, followed by a standard second dose, the efficacy shot up to 90%. This was a very unexpected yet interesting finding, which influenced the modification of future vaccination schedules of this vaccine31. Subsequently, the pooled primary analysis of the data confirmed the findings of the two-dose regimen reported in the interim analysis. The findings reaffirmed that the vaccine was highly efficacious and the efficacy was related to the dosing interval employed. This study suggested that an interval of up to 12 weeks could be advantageous over 4 weeks, as initially suggested. The authors argued that this longer interval between the two doses would enable the vaccination of a greater number of people, considering the global shortage of the vaccine32.
Sputnik V vaccine: This is the Russian vaccine, developed by the Gamaleya Research Institute of Epidemiology and Microbiology with funding from the Russian Direct Investment Fund (RDIF). This vaccine is technically termed Gam-COVID-Vac, but is more widely known as Sputnik V after the series of famous Russian spacecraft. This is a two-dose vaccine that is administered 21 days apart. It uses a non-replicating adenovirus vector, just like the Oxford University/AstraZeneca vaccine. However, two different types of adenoviruses are used: Ad26 for the first dose and Ad5 for the second dose, which expresses the full-length S protein of SARS-CoV-2. In two open, non-randomized Phase 1/2 clinical trials, it was shown to be safe and induced strong humoral and cellular immune responses. The binding antibodies were ten-fold higher in the vaccine group, compared to the convalescent group. However, NAb levels were not significantly different in either of the groups. Regarding cell-mediated immunity, the proliferation of CD4+ and CD8+ T-cells and production of IFN-γ were observed in PBMCs, when they were stimulated with the antigen33. A subsequent Phase 3, randomized, double-blind, placebo-controlled clinical trial conducted at 25 healthcare facilities in Moscow, showed that Sputnik V exhibited an efficacy of 91.6% following the second dose34.
A variation of Sputnik V is the so-called Sputnik Light, which is a single-dose vaccine. In this vaccine, essentially the first dose (Ad26) of Sputnik V is administered and has exhibited an efficacy of 79.4% in Phase 1/2 clinical trials in Russia.
Janssen vaccine: This is a single-dose vaccine developed by Janssen, which is owned by the American Pharmaceutical Giant, Johnson & Johnson. As of August 31st, 2021 it was the only single-dose vaccine to have received EUL approval by WHO. It is a non-replicating viral vector vaccine that uses Ad26 as the vector to carry the transgene encoding the full-length S protein of SARS-CoV-2. That is why it is technically termed Ad26.COV2.S. In Phase 3 trials, it exhibited an efficacy of 67%35. A randomized, multicentre, placebo-controlled, Phase 1-2a clinical trial of Ad26.COV2.S was conducted in individuals aged 18-55 years (cohort 1) and those 65 years of age or above (cohort 2). A single-dose or two-dose schedule was followed. NAbs were detected in over 90% of vaccinees on day 29 after the first dose (GMT: 212-354) and reached 96% by day 57 with a further increase in the NAb titers (GMT: 288-488). These titers remained stable till day 71. Following the second dose, the titers increased by 2.6 to 2.9 folds. The binding antibody levels were similar to the NAb levels. On day 15 after the second dose, the CD4+ T-cell responses were detected in 60-83% of individuals, depending on the cohort they belonged to. These CD4+ T-cell responses primarily exhibited a Th1 polarization. Overall, the CD8+ T-cell responses were very robust36.
Inactivated vaccines: Inactivated vaccines, also known as killed vaccines, are so-called because the viral particles are inactivated using chemical treatments, such as β-propiolactone. Hence, they don’t pose any threat of causing infection or reversion of infectivity. Inactivated vaccines, along with live-attenuated vaccines (LAV), are developed using time-tested techniques, which have been around for over a century. Hence, these are sometimes referred to as traditional vaccines. There are currently 16 inactivated candidate vaccines in clinical trials. The major inactivated vaccines have been discussed below.
Sinovac vaccine: This is a two-dose vaccine that is administered 14-28 days apart. It is a Chinese vaccine, developed by Sinovac Biotech and named CoronaVac. It is developed by growing SARS-CoV-2 (CN02 strain) in cell culture using African green monkey kidney cells, popularly known as the Vero cell line. After the culturing process is over, the viral particles are harvested, inactivated with β-propiolactone, concentrated and purified. The vaccine is then adsorbed to the adjuvant aluminium hydroxide before immunization. The whole process is conducted under stringent Good Manufacturing Practices (GMP) conditions37.
A randomized, double-blind, placebo-controlled Phase 1/2 clinical trial of CoronaVac was conducted in the 18-59 years age group, using two doses (3 μg and 6 μg). In the Phase 1 trial, NAbs were detected after the second dose on day 28 in 83% of individuals in the 3 μg group and 79% in the 6 μg group. In the Phase 2 trial, NAbs were detected after the second dose on day 28 in 97% of individuals in the 3 μg group and 100% in the 6 μg group. Taking into consideration safety and immunogenicity, the study findings suggested that the 3 μg dose of CoronaVac would be most suitable for testing in Phase 3 clinical trials37. Similar conclusions were reached when the same study was conducted in the age group of 60 years and above38.
A Phase 3 clinical trial of CoronaVac in Turkey, conducted on 6,648 individuals showed an efficacy of 83.5%. However, the same vaccine showed an efficacy of only 50.4% in a Phase 3 clinical trial conducted in Brazil. However, the Brazilian researchers indicated that this efficacy value was for patients with very mild diseases who didn’t require medical assistance. They further added that the vaccine exhibited an efficacy of 78% in mild cases who needed medical assistance and 100% in those with moderate to severe disease. Surprisingly, Indonesia reported an efficacy of 65.3%. Hence, there is some dilemma about the efficacy of this vaccine as it performed differently in different countries39,40. As of August 2021, there was no information on the cellular immune responses of this vaccine in humans.
Sinopharm vaccine: Like CoronaVac, this is also a two-dose vaccine, which has been named BBIBP-CorV. This vaccine is administered 21 days apart and is produced in exactly the same way as CoronaVac. A randomized, double-blind, placebo-controlled, Phase 1/2 clinical trial was conducted with BBIBP-CorV. The vaccine was found to be safe and well-tolerated at all doses tested in the two age groups (18-59 years and >60 years). Binding antibodies were detected in all vaccines by day 42. Concerning NAb inductive potency, a two-dose vaccination schedule (0, 21 days or 0, 28 days) at a dose of 4 μg was found to be far superior to other doses and schedules41. A Phase 3 clinical trial of BBIBP-CorV conducted on 31,000 individuals in Dubai showed that the vaccine had an efficacy of 86%. There was a 99% seroconversion rate in the case of NAbs and 100% effectiveness in preventing moderate to severe disease. Moreover, there were no safety concerns with the vaccine. As of August 2021, no information was available on the cellular immune responses of this vaccine in humans42.
Live-attenuated vaccines: Unlike inactivated vaccines, in the case of live-attenuated vaccines (LAV), the virus is not killed but is weakened by chemical treatments. A fine balance has to be struck during attenuation so that the virus is weakened to such an extent that it loses its virulence or infectivity, but still retains its immunogenicity i.e., the ability to stimulate the immune system to generate an immune response. There are currently only two LAV candidates undergoing clinical development. These have been developed by Codagenix and Meissa. The former is slightly more advanced and is undergoing Phase 1/2 clinical trials, while the latter is still in Phase 1. Codagenix’s COVI-VAC vaccine is discussed below.
The COVI-VAC is an intranasal vaccine that can be used as a single-dose or two-dose regimen, administered 28 days apart. The vaccine uses Codagenix’s proprietary deoptimization technology and is safe and effective in animal models even after a single dose and is currently being evaluated in Phase 1/2 clinical trials. The COVI-VAC generates antibodies, not just against the S protein, but also against the other two surface proteins, namely, E protein and M protein, thereby being polyspecific. It elicits robust humoral and cell-mediated immune responses43.
A randomized, double-blind, placebo-controlled, dose-escalation Phase 1 clinical trial of COVI-VAC is currently ongoing and is scheduled to be completed in May 2022. It is the first study of the vaccine in humans. It is evaluating the safety and immunogenicity of the vaccine in 48 individuals aged 18-30 years. The individuals were divided into three groups, who received either low, medium or high doses of the vaccine or a placebo (saline), administered intranasally. Results of this study are not yet available44.
A randomized, placebo-controlled, observer-blind Phase 1/2 clinical trial of COVI-VAC is currently being conducted in Vietnam. This prospective, single-centre study aims to assess the safety and immunogenicity of the vaccine in 420 individuals aged 18-75 years. The trial is scheduled to be completed on September 30th, 2022. Like the above Phase 1 study, data for this study is also not yet available45.
Virus-like particle vaccines: As the name suggests, a virus-like particle (VLP) is like a virus, but not exactly so. Unlike a virus, it doesn’t contain any genetic material (RNA in the case of SARS-CoV-2), which makes it non-infectious and therefore, very safe. So, it resembles a football, having a shell, without having anything inside. In VLPs, the surface proteins remain intact. Hence, all three surface structural proteins of SARS-CoV-2, namely, S, E and M proteins all remain unaltered. This allows the VLP to stimulate the immune system, without the risk of infection. These structural proteins are expressed in various living or cell-free expression systems, followed by self-assembly, which gives shape and imparts structural integrity to the VLP46.
There are currently five VLP-based candidate vaccines in clinical trials. The VLPs are highly immunogenic and generate robust humoral and cell-mediated immune responses13.
Comparing the immunogenicity of different candidate vaccines is difficult: Comparing the immunogenicity of different vaccines is often a difficult and futile exercise, as the assays used in different studies are not always similar and therefore, incomparable. For example, ELISA titers vary in-between studies, as some measure end-point titers, while others measure mid-point titers, making it difficult to interpret the findings. The same thing often happens with PRNT50 assays, as some in vitro assays use pseudoviruses while others use live viruses. Moreover, antibody titers in assays are often compared with those present in convalescent sera, which is also likely to be fallacious, as antibody titers generated by natural infections would vary between individuals15.
Adjuvants play a key role in boosting the immune response: Adjuvants are chemical substances added to vaccines to enhance the magnitude and durability of antigen-induced immune responses. Adjuvants can stimulate various pathways associated with innate immunity, as well as skew the humoral and cell-mediated immune responses toward the desired direction. Some of the common adjuvants currently being used in the COVID-19 vaccines include aluminium hydroxide, MF59 (oil-in emulsion), Toll-like receptor (TLR) agonists (e.g., CpG1018) and the adjuvant system (AS) series (e.g., AS03) developed by GlaxoSmithKline (GSK). In this context, the Matrix-MTM adjuvant has already been discussed above in the context of the Novavax vaccine47.
Intranasal vaccine
A promising new technology for generating mucosal immunity: Besides COVI-VAC, the intranasal Codagenix vaccine that belongs to the LAV category, there is another intranasal vaccine currently under development, which is an adenoviral-vectored vaccine. A randomized, single-centre, open-label and Phase 1 clinical trial was recently conducted in Wuhan, China, where aerosolized Ad5-nCoV was delivered intranasally. This procedure was simple, painless, well-tolerated and generated a robust immune response, including induction of NAbs. This candidate vaccine will proceed to Phase 2/3 clinical trials very soon48.
Intranasal vaccines are advantageous over other vaccines because they can produce IgA antibodies that generate mucosal immunity. Since the nasal cavity is the major portal of entry of SARS-CoV-2, stimulation of the nasal mucosa to generate IgAs would neutralize the virus and prevent it from entering the lungs and beyond. The feasibility of this approach is exemplified by the fact that IgA molecules have been detected in the nasal washings of convalescent patients and these have been found to neutralize SARS-CoV-249.
CONCLUSION
Unprecedented progress has been made on the vaccine front in the past two years. Much has been learned about the immune effector mechanisms, including the role of binding antibodies, NAbs, T-cells and innate immune responses, in the context of our fight against the virus. Elucidating these protective mechanisms, as well as identifying the correlates of protection are of the utmost importance in furthering our efforts to not only develop other novel vaccines but also tweak the existing ones in the light of the emergence of variants worldwide. Hence, the battle is still on between the vaccines and the variants. Only time will tell who will win.
SIGNIFICANCE STATEMENT
This study explores the characteristics of the various COVID-19 vaccine platforms, with special reference to the immune responses that they generate. This aspect is often neglected in the medical literature, but is nevertheless, very crucial and the review fills the knowledge gaps in this area. The importance of the review lies in the fact that knowing the type of immune responses generated by the currently available vaccines, will help to design future candidate vaccines that can be tweaked to generate targeted immune responses that specifically neutralize the emerging variants that exhibit the phenomenon of “immune escape”. This is likely to be a game-changer in the fight against COVID-19 and will have an immense social impact as it will be instrumental in eventually ending the pandemic.
REFERENCES
- O’Driscoll, M., G.R.D. Santos, L. Wang, D.A.T. Cummings and A.S. Azman et al., 2021. Age-specific mortality and immunity patterns of SARS-CoV-2. Nature, 590: 140-145.
- Diamond, M.S. and T.C. Pierson, 2020. The challenges of vaccine development against a new virus during a pandemic. Cell Host Microbe, 27: 699-703.
- Bharati, K., R. Rani and S. Vrati, 2009. Evaluation of Japanese encephalitis virus DNA vaccine candidates in rhesus monkeys [Macaca mulatta]. Vaccine, 27: 10-16.
- Bharati, K. and S. Vrati, 2012. Viral vaccines in India: An overview. Proc. Natl. Acad. Sci. India Sect. B: Biol. Sci., 82: 181-198.
- Wu, F., S. Zhao, B. Yu, Y.M. Chen and W. Wang et al., 2020. A new coronavirus associated with human respiratory disease in China. Nature, 579: 265-269.
- Benvenuto, D., M. Giovanetti, A. Ciccozzi, S. Spoto, S. Angeletti and M. Ciccozzi, 2020. The 2019‐new coronavirus epidemic: evidence for virus evolution. J. Med. Virol., 92: 455-459.
- Forni, D., R. Cagliani, M. Clerici and M. Sironi, 2017. Molecular evolution of human coronavirus genomes. Trends Microbiol., 25: 35-48.
- Chen, Y., Q. Liu and D. Guo, 2020. Emerging coronaviruses: genome structure, replication, and pathogenesis. J. Med. Virol., 92: 418-423.
- Hoffmann, M., H. Kleine-Weber, S. Schroeder, N. Krüger and T. Herrler et al., 2020. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell, 181: 271-280.
- Lu, R., X. Zhao, J. Li, P. Niu and B. Yang et al., 2020. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet, 395: 565-574.
- Ahmed, S.F., A.A. Quadeer and M.R. McKay, 2020. Preliminary identification of potential vaccine targets for the COVID-19 coronavirus (SARS-CoV-2) based on SARS-CoV immunological studies. Viruses, Vol. 12(3), 254.
- Mukherjee, S., D. Tworowski, R. Detroja, S.B. Mukherjee and M. Frenkel-Morgenstern, 2020. Immunoinformatics and structural analysis for identification of immunodominant Epitopes in SARS-CoV-2 as potential vaccine targets. Vaccines, Vol.8(2), 290.
- Zieneldien, T., J. Kim, J. Cao and C. Cao, 2021. COVID-19 vaccines: Current conditions and future prospects. Biology, Vol. 10(10), 960.
- Bharati, K., 2020. COVID-19 pandemic: Will a vaccine be available soon? J. Clin. Diagn. Res., 14: AB01-AB03.
- Grigoryan, L. and B. Pulendran, 2020. The immunology of SARS-CoV-2 infections and vaccines. Semin. Immunol., Vol. 50, 101422.
- Alka, K. Bharati, Y.P.S. Malik and S. Vrati, 2007. Immunogenicity and protective efficacy of the E. coli-expressed domain III of Japanese encephalitis virus envelope protein in mice. Med. Microbiol. Immunol., 196: 227-231.
- Keech, C., G. Albert, I. Cho, A. Robertson, P. Reed and S. Nea et al., 2020. Phase 1–2 trial of a SARS-CoV-2 recombinant spike protein nanoparticle vaccine. N. Engl. J. Med., 383: 2320-2332.
- Heath, P.T., E.P. Galiza, D.N. Baxter, M. Boffito and D. Browne et al., 2021. Safety and efficacy of NVX-CoV2373 Covid-19 vaccine. N. Engl. J. Med., 385: 1172-1183.
- Walsh, E.E., R.W. Frenck, A.R. Falsey, N. Kitchin and J. Absalon et al., 2020. Safety and immunogenicity of two RNA-Based Covid-19 vaccine candidates. N. Engl. J.Med., 383: 2439-2450.
- Chagla, Z., 2021. The BNT162b2 (BioNTech/Pfizer) vaccine had 95% efficacy against COVID-19 ≥7 days after the 2nd dose. Ann. Intern. Med. Vol. 174, No. 2.
- Sahin, U., A. Muik, E. Derhovanessian, I. Vogler and L.M. Kranz et al., 2020. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature, 586: 594-599.
- Baden, L.R., H.M. El Sahly, B. Essink, K. Kotloff and S. Frey et al., 2020. Efficacy and safety of the mRNA-1273 SARS-CoV-2 vaccine. N. Engl. J. Med., 384: 403-416.
- Jackson, L.A., E.J. Anderson, N.G. Rouphael, P.C. Roberts and M. Makhene et al., 2020. An mRNA vaccine against SARS-CoV-2-preliminary report. N. Engl. J. Med., 383: 1920-1931.
- Widge, A.T., N.G. Rouphael, L.A. Jackson, E.J. Anderson and P.C. Roberts et al., 2020. Durability of responses after SARS-CoV-2 mRNA-1273 vaccination. N. Engl. J. Med., 384: 80-82.
- Bharati, K. and S. Vrati, 2013. DNA vaccines: Getting closer to becoming a reality. Indian J. Med. Res., 137: 1027-1028.
- Bharati, K., M.B. Appaiahgari and S. Vrati, 2005. Effect of cytokine-encoding plasmid delivery on immune response to Japanese encephalitis virus DNA vaccine in mice. Microbiol. Immunol., 49: 349-353.
- Silveira, M.M., G.M.S.G. Moreira and M. Mendonça, 2020. DNA vaccines against COVID-19: Perspectives and challenges. Life Sci., Vol. 267, 118919.
- Khobragade, A., S. Bhate, V. Ramaiah, S. Deshpande and K. Giri et al., 2022. Efficacy, safety, and immunogenicity of the DNA SARS-CoV-2 vaccine (ZyCoV-D): The interim efficacy results of a phase 3, randomised, double-blind, placebo-controlled study in India. Lancet, 399: 1313-1321.
- Smith, T.R.F., A. Patel, S. Ramos, D. Elwood and X. Zhu et al., 2020. Immunogenicity of a DNA vaccine candidate for COVID-19. Nat. Commun., Vol, 11: 2601.
- Folegatti, P.M., K.J. Ewer, P.K. Aley, B. Angus and S. Becker, 2020. Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: A preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet, 396: 467-478.
- Voysey, M., S.A.C. Clemens, S.A. Madhi, L.Y. Weckx and P.M. Folegatti et al., 2021. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet, 397: 99-111.
- Voysey, M., S.A.C. Clemens, S.A Madhi, L.Y Weckx and P.M Folegatti et al., 2021. Single-dose administration and the influence of the timing of the booster dose on immunogenicity and efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine: A pooled analysis of four randomised trials. Lancet, 397: 881-891.
- Logunov, D.Y., I.V. Dolzhikova, O.V. Zubkova, A.I. Tukhvatulin and D.V. Shcheblyakov et al., 2020. Safety and immunogenicity of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine in two formulations: Two open, non-randomised phase 1/2 studies from Russia. Lancet, 396: 887-897.
- Logunov, D.Y., I.V. Dolzhikova, D.V. Shcheblyakov, A.I. Tukhvatulin and O.V. Zubkova et al., 2021. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: An interim analysis of a randomised controlled phase 3 trial in Russia. Lancet, 397: 671-681.
- Stephenson, K.E., M.L. Gars, J. Sadoff, A.M. de Groot and D. Heerwegh et al., 2021. Immunogenicity of the Ad26.COV2.S vaccine for COVID-19. JAMA, 325: 1535-1544.
- Sadoff, J., M. le Gars, G. Shukarev, D. Heerwegh and C. Truyers et al., 2021. Interim results of a phase 1-2a trial of Ad26.COV2.S Covid-19 vaccine. N. Engl. J. Med., 384: 1824-1835.
- Zhang, Y., G. Zeng, H. Pan, C. Li and Y. Hu et al., 2020. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18–59 years: A randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect. Dis., 21: 181-192.
- Wu, Z., Y. Hu, M. Xu, Z. Chen and W. Yang et al., 2021. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine (CoronaVac) in healthy adults aged 60 years and older: A randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect. Dis., 21: 803-812.
- Palacios, R., E.G. Patiño, R. de Oliveira Piorelli, M.T.R.P. Conde and A.P. Batista et al., 2020. Double-blind, randomized, placebo-controlled phase III clinical trial to evaluate the efficacy and safety of treating healthcare professionals with the adsorbed COVID-19 (Inactivated) vaccine manufactured by Sinovac – PROFISCOV: A structured summary of a study protocol for a randomised controlled trial. Trials, Vol. 21: 853.
- Akova, M. and S. Unal, 2021. A randomized, double-blind, placebo-controlled phase III clinical trial to evaluate the efficacy and safety of SARS-CoV-2 vaccine (inactivated, Vero cell): A structured summary of a study protocol for a randomised controlled trial. Trials, Vol. 22: 276.
- Xia, S., Y. Zhang, Y. Wang, H. Wang and Y. Yang et al., 2021. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBIBP-CorV: A randomised, double-blind, placebo-controlled, phase 1/2 trial. Lancet Infect. Dis., 21: 39-51.
- Kandeil, A., A. Mostafa, R.R. Hegazy, R. El-Shesheny and A. El-Taweel et al., 2021. Immunogenicity and safety of an Inactivated SARS-CoV-2 vaccine: Preclinical studies. Vaccines, Vol. 9(3), 214.
- Wang, Y., C. Yang, Y. Song, J.R. Coleman and M. Stawowczyk et al., 2021. Scalable live-attenuated SARS-CoV-2 vaccine candidate demonstrates preclinical safety and efficacy. Proc. Natl. Acad. Sci., Vol, 118, No. 29, e2102775118.
- Tasker, S., D. Bendel, M. Bevan, S. Mueller, A. Kushnir, B. Londt and J.R. Coleman, 2021. 584. Phase 1 placebo-controlled trial of COVI-VAC™, an intranasal, live attenuated COVID-19 vaccine. Open Forum Infect. Dis., 8: S394-S394.
- Alu, A., L. Chen, H. Lei, Y. Wei, X. Tian and X. Wei, 2022. Intranasal COVID-19 vaccines: From bench to bed. eBioMed. Vol, 76: 103841.
- Nooraei, S., H. Bahrulolum, Z.S. Hoseini, C. Katalani, A. Hajizade, A.J. Easton and G. Ahmadian, 2021. Virus-like particles: preparation, immunogenicity and their roles as nanovaccines and drug nanocarriers. J. Nanobiotech., Vol. 19, 59.
- Dong, Y., T. Dai, Y. Wei, L. Zhang, M. Zheng and F. Zhou, 2020. A systematic review of SARS-CoV-2 vaccine candidates. Signal Transduction Targeted Ther., Vol. 5, 237.
- Wu, S., J. Huang, Z. Zhang, J. Wu and J. Zhang et al., 2021. Safety, tolerability, and immunogenicity of an aerosolised adenovirus type-5 vector-based COVID-19 vaccine (Ad5-nCoV) in adults: Preliminary report of an open-label and randomised phase 1 clinical trial. Lancet Infect. Dis., 21: 1654-1664.
- Butler, S.E., A.R. Crowley, H. Natarajan, S. Xu and J.A. Weiner et al., 2021. Distinct features and functions of systemic and mucosal humoral immunity among SARS-CoV-2 convalescent individuals. Front. Immunol., Vol. 11: 618685.
How to Cite this paper?
APA-7 Style
Bharati,
K. (2023). Immune Responses Elicited by COVID-19 Vaccines. Asian Journal of Biological Sciences, 16(1), 89-102. https://doi.org/10.3923/ajbs.2023.89.102
ACS Style
Bharati,
K. Immune Responses Elicited by COVID-19 Vaccines. Asian J. Biol. Sci 2023, 16, 89-102. https://doi.org/10.3923/ajbs.2023.89.102
AMA Style
Bharati
K. Immune Responses Elicited by COVID-19 Vaccines. Asian Journal of Biological Sciences. 2023; 16(1): 89-102. https://doi.org/10.3923/ajbs.2023.89.102
Chicago/Turabian Style
Bharati, Kaushik.
2023. "Immune Responses Elicited by COVID-19 Vaccines" Asian Journal of Biological Sciences 16, no. 1: 89-102. https://doi.org/10.3923/ajbs.2023.89.102

This work is licensed under a Creative Commons Attribution 4.0 International License.