Trichoderma longibrachiatum’s Secondary Metabolite-A Review on Potential Compounds for Plant Growth and Biological Control for Plant Pathogens
| Received 14 Dec, 2022 |
Accepted 27 Jun, 2023 |
Published 30 Sep, 2023 |
Indeed, a significant amount of agricultural production has been lost due to plant diseases that were caused by phytopathogens. Chemical pesticides are applied to protect the crops from the reduce substantial yield loss. Improved crop yields have been achieved for a long time through the use of synthetic chemical pesticides. However, the use of these pesticides may one day be limited due to their negative effects on human health and the environment. Global ecological awareness of the use of natural products and microorganisms to manage plant diseases has led to the use of beneficial antagonistic bacteria and fungi in different methods. Several microorganisms limit pathogen growth or indirectly increase plant-mediated resistance. Trichoderma is one of the most effective biological control agents for soil and foliar diseases. The Trichoderma spp., biocontrol potential depends on number of mechanisms such as antibiosis, mycoparasitism and the host induced systemic resistance. As typically recognised saprophytic fungi, Trichoderma species generate and exude a wide range of secondary metabolites into their environment while having little nutritional requirements. The non-ribosomal peptides (peptobiols, siderophores, gliotoxin and glovirin), polyketides, terpenes, pyrones and isocyanine are some of the Trichoderma spp., derived secondary metabolites. These metabolites are associated in different biological activities like biocontrol activities and or microbial intrractions. Since, it’s becoming more important to extract these molecules from safer, biodegradable antifungal solutions, which may be the next generation of biological pesticides, studies on Trichoderma’s antifungal active components are intensifying. This article is reviewed particularly about the major secondary metabolites that are produced by the beneficial fungus Trichoderma longibrachiatum and enhance the present knowledge on the potential compounds for plants.
| Copyright © 2023 Ramasamy et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Phytopathogens cause major damage and losses, affecting agricultural production. Chemical fungicides, disease-resistant cultivars and other cultural practices are used to manage fungal plant diseases. Overuse of chemical fungicides directly threatens human, animal and environmental health. Because pesticides have the potential to affect the microbial communities linked with plants in the soil, it is essential to look for environmentally acceptable alternatives. Decades have passed since the invention of microbiological tools and biocontrol agents, such as bacteria and fungi as biocontrol agents that are microbial inoculants to replace insecticides. Live organisms suppress plant pathogen populations through biological control. A microbe’s ability to create antibiotics or enzymes that lyse microbial cell walls, deplete iron from the nearby plant root system, develop disease resistance or compete with pathogens for niches in the rhizosphere are all examples of how they can exert biocontrol. Because they can boost crop growth and the availability and uptake of nutrients. The microbial based biocontrol agents are tools that hold great promise for the development of sustainable agriculture. Microbial biological control agents (MBCAs) are applied to crops to manage plant diseases. Products containing Trichoderma spp., are widely used because these fungi reduce disease and nematode population on host plants soils, mitigating abiotic stress. The Trichoderma sp. are being antagonistic towards Fusarium sp., Pythium sp. and Sclerotium rolfsii, Rhizoctonia sp. and also has the ability to suppress the growth of wood-rotting fungus, including Botrytis sp., Verticillium sp., Sclerotia sp. and Gaeumannomyces sp. Secondary metabolites of mycoparasitic fungi will contribute to the discovery of new antagonistic substances in the future. The most varied group of phytopathogens, fungi, have a significant impact on agriculture. The pathogenic fungus can become resistant to fungicides and infect new hosts due to their extensive genetic flexibility and broad-spectrum lifecycles. Resistance to compounds that have already been applied is a growing concern in agriculture, so there is an urgent need for the discovery of environmentally sustainable new antagonistic substances for controlling diseases. As a result, new management techniques are required to combat pathogenic fungi. Secondary metabolites from Trichoderma spp., have been used to protect plants from a number of phytopathogens in an efficient and effective manner. The phytopathogenic secondary metabolites from Trichoderma longibrachiatum were presented in this review. This article has covered a few aspects of the structural overview of secondary metabolites and the biogenesis of these molecules and also discussed overview of the secondary metabolites that are produced by Trichoderma longibrachiatum.
Mycoparasitic fungi: In order to improve their fitness and chances of survival, mycoparasitic fungi that feed on other fungi produce both volatile and non-volatile secondary metabolites. The excretion of siderophores, which result in high affinity iron chelation, is significantly upregulated in iron-limiting situations. Several antimicrobial metabolites also support in the persistence of ecological niches1,2. Due to the mycoparasitic lifestyle, there is substantial excretion of secondary metabolites and overrepresentation of genes related to secondary metabolism, which enables the fungus to access its prey and either survive in or kill the host3.
Additionally, certain fungal secondary metabolites have been shown to have positive effects on plants, including promoting root and shoot vitality and growth, enhancing the plant’s resistance to abiotic stresses and activating its immune system are both outcomes of this process (induced systemic resistance, ISR) to enhance resistance and chances of being able to survive the threat of possible diseases caused by pathogens4. Evidence has recently accumulated that several secondary metabolites also function as molecules of communication across species boundaries5-6. The fungi kingdom is home to a wide variety of mycoparasitic species, particularly those belonging to the order Hypocreales7. Trichoderma (teleomorph hypocrea), a widespread, diversified fungus genus, is one of the most well-studied instances in this regard3. Necrotrophic mycoparasitic species of Trichoderma are used successfully in agriculture as biocontrol agents against agricultural plant pathogenic fungus. In addition to this, it is considered that they improve the immune systems of plants and encourage growth, vigour and resistance across the entire system. Because different species, ecological conditions and each strain of Trichoderma can give rise to a diverse set of chemical byproducts (Fig. 1), there is reason to be optimistic about the possibility of the discovering novel compounds1,3.

|
Secondary metabolites: Microorganism-derived secondary metabolites (SMs) may have an antifungal effect on economically significant phytopathogenic fungi8. Fungi are responsible for the production of 42% of the approximately 23,000 identified microbial secondary metabolites that serve as important sources for medicines with therapeutic applications9,10. Researchers have shown that fungus and fungal metabolites may be useful in preventing and treating plant illnesses brought on by fungal pathogens. Trichoderma spp., are among the fungi that have received the most attention from researchers and are now used in clinical settings due of the diverse spectrum of antimicrobial SMs that they produce11,12. Trichoderma species produce a multitude of metabolites that are then secreted into their environment while having very few dietary requirements. This review article recognized the value of these metabolites due to their potential use in agriculture, industry and medicine. Trichoderma spp., despite their low dietary demands, produce an abundance of secondary metabolites (SMs) that have antifungal effects against phytopathogenic fungi. These SMs include terpenes, pyrones, gliotoxin, gliovirin and peptaibols11,13,14. Various secondary metabolites, including virdin, harzianolde, glioviridin and growth promoting inducers (IAA, auxins, etc.), have been linked to the success of Trichoderma species as plant growth promoters15. Other fungus produces useful VOCs through various methods that influence biological regulation, trigger defense reactions and stimulate plant growth16.
Trichoderma longibrachiatum: A member of the anamorphic genus Trichoderma, clade Longibrachiatum, order Sordariomycetes, family Hypocreaceae and phylum Sordariomycetes, Trichoderma longibrachetum is found among the hypocrealean fungi21. There have been reports of several strains of T. longibrachiatum that show promise as possible biocontrol agents against plant diseases caused by fungi, bacteria and nematodes17,21-25. T. longibrachiatum has been used to get proteins, quinones and triterpenoids, which are all new antibiotics. This resulted in the realization that T. longibrachiatum could be a source of new antibiotics (Table 1)17-20,26. One of the most recent biocontrol fungi to be examined is Trichoderma longibrachiatum. A number of new volatile and nonvolatile metabolites, including proteins, quinones and hydrocarbons, have so far been discovered in different strains. Due to this,
| Table 1: | Secondary metabolites secreted by Trichoderma species | |||
| Compound | Source | Activity | References |
| Trichothecinol A, | Isolated from the root of | Potent antifungal activity | Du et al.17 |
| 8-deoxy-trichothecin, trichothecinol B | Suaeda glauca, a highly | Moderate nematicidal activity | |
| 10-cycloneren-3,5,7-triol, | halophile plant | Potent antifungal activity | |
| 10(E)-cyclonerotriol | |||
| Trichodermene A | |||
| Deoxytrichodermaerin | Isolated from marine red alga L. okamurai |
Potent antimicroalgal activity | Zou et al.18 |
| Homodestcardin, | Isolated from the root | Moderate nematicidal activity | Du et al.17 |
| trichomide B, homodestruxin B | of Suaeda glauca | ||
| Peptide-Trichokonin VI (Tk VI) | Inhibits primary root growth in A. thaliana (plant-microbe interactions) |
Shi et al.19 | |
| Mycoparasitism-related metabolites | Antifungal properties via inhibition | ||
| Bisorbicillinoid-Bisvertinolone | of β-(1,6)-glucan biosynthesis | Kontani et al.20 |
T. longibrachiatum has gained recognition as a potential source of novel antibiotics and as a helpful biocontrol agent against nematodes and bacteria that harm plants24,27. The growth of the pathogens was slowed down by these metabolites. Trichoderma longibrachiatum’s secondary metabolites, for instance, have been shown to be effective against Candida albicans and Pyricularia oryzae28. In addition to this, it was discovered that the secondary metabolites could stimulate protection against disease and modulate the growth of plants16.
Trichoderma longibrachiatum is a fungus that is typically isolated from terrestrial soil and plants. It has a wide range of potential antagonistic effects on plant pathogenic fungi, which can impede their growth. Regarding the antifungal substances obtained from T. longibrachiatum, they shown a strong antagonistic potential against the strains of Botrytis cinerea that are carbendazim-resistant, Colletotrichum lagenarium, C. fragariae and Fusarium spp.17,29-31.
The metabolites of T. longibrachiatum were recently found to have antifungal effects against two important pathogens Sclerotium rolfsii and Macrophomina phaseolina. Many antifungal substances were released by T. longibrachiatum upon direct interactions with pathogens, (longifolene, 1-butanol 2-methyl, cedrene, caryophyllene and cuprenene). They function as intermediates in the biosynthesis of sesquiterpenoids, alkanes and the degradation of trimethylamine. Even more so, 1-pentanol, 1-hexanol, myristonyl-pantothenate, bisabolol, d-Alanine and diethyl trisulphide, attracts interest as a potentially effective antibacterial chemical with the added benefit of promoting plant growth16.
Multiple volatile organic compounds (VOCs) were detected in T. longibrachiatum, including monomers, esters, ketones, alkynes, esters, enzymes, carotenoids and a camphene derivative. Here, among the bioactive VOCs, epi-caryophyllene was identified as having antifungal and antioxidant activities, along with boosting seedling development & chlorophyll concentration32,33.
Epipolythiodioxopiperazines: Among secondary metabolites of fungi, epipolythiodioxopiperazines (ETPs) are distinguished by a diketopiperazine ring that derives from a peptide and have a high reactive potential. Since they are produced from protein hydrolysates, diketopiperazines (DKPs) are regarded as the byproduct of protein degradation and are often not desirable peptides. Epipolythiodioxopiperazines (ETPs) are toxic because they bind to proteins via disulphide bridges that generate reactive oxygen species via redox cycles, so rendering proteins inert34.
Because of their potent biological activities, scientists have focused their efforts increasingly towards DKP study in recent years. Numerous DKPs derived from microbes were identified and their biological functions investigated.
|
|

|
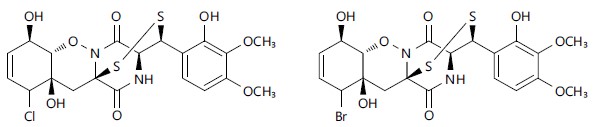
The class of toxins known as epipolythiodioxopiperazines includes both gliovirin and gliotoxin, which share the characteristic disulphide bridges35.
Trichoderma longibrachiatum produced two gliovirin analogues (Fig. 2) which showed antifungal properties against R. Solani11.
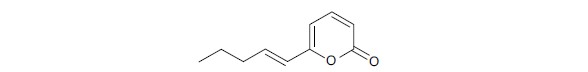
Pyrones: The flavouring ingredient that gives coconut its distinctive aroma is called 6-pentyl-2H-pyran-2-one (6-PP) and studies have shown that it possesses antifungal and plant growth-promoting properties6. It belongs to the large class of molecules known as volatile organic compounds, which include many small molecule metabolites that have a high vapour pressure at room temperature and a low solubility in water (VOCs)36. A pyrone analogue (Fig. 3) was acquired from the fungi Trichoderma harzianum and T. longibrachiatum possess antimicrobial effects towards Armillaria melanoma37.
It was proposed that the synthesis of pyrones was caused by the oxidation of linoleic acid to 13-hydroperoxide-diene, followed by the formation of 5-hydroxy-2,4-decenioc acid and esterification38.
Butenolides: The 5-Hydroxyvertinolide (Fig. 4), an antifungal butenolide from the fungus T. longibrachiatum, was discovered to be antagonistic to the fungus Mycenacitricolor, the cause of coffee’s American leaf spot disease39. Most likely, two Favorskii rearrangements of a C-14-diepoxide are involved in the biosynthesis of these butenolides, with the two carbons that make up the lactone being driven out at the end.
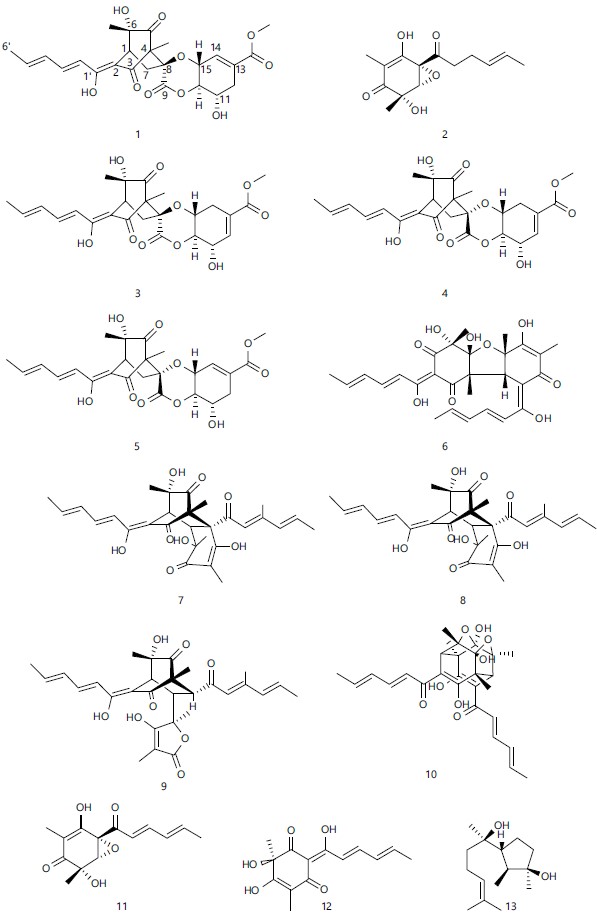
Sorbicillinoids: A family of hexaketide fungal metabolites known as sorbicillinoids has a conventional or altered sorbyl side chain. The first found sorbicillin was in Penicillium notatum in 1948 as an impurity/contaminant and later in aquatic and terrestrial microbes found more than 100 sorbicillinoids (Acremonium, Aspergillus, Clonostachys, Eurotiomycete, Penicillium, Phaeoacremonium, Phialocephala, Paecilomyces, Scytalidium, Trichoderma and Verticillium)40-42.
|
|
The structural characteristics of different sorbicillinoids allow them to be divided into monomers, dimers, trimers and hybrids43,44.
There are 12 different sorbicillinoids in this study (Fig. 5), but they can be put into three different groups: Monomeric sorbicillinoids, dimeric sorbicillinoids and hybrid sorbicillinoids. Bisvertinolone, which is the strongest antifungal agent, is thought to be made in the body from oxosorbicillinol and sorbicillinol through a Michael-type reaction45. The investigated phytopathogenic fungi responded most favourably to the antifungal agents oxosorbicillinol, bisvertinolone and epoxysorbicillinol. Similar to this, it has been noted that bisvertinolone (6) inhibits b-1,6-glucan production in developing hyphal cells of P. capsici, causing morphological malformations to occur20. In this work, oxosorbicillinol (12) demonstrated a moderate antimicrobial activity against plant fungi such Colletotrichum coccodes, Magnaporthe oryzae and late blight causing fungus Phytophthora infestans46, with MIC values ranging from 25 to 400 g mL–1.
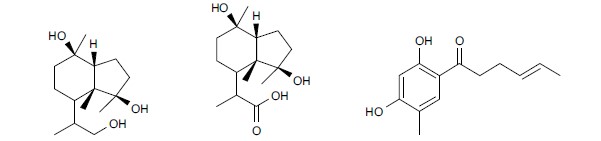
Other compounds: Three antifungal compounds were obtained from Trichoderma longibrachiatum, 10, 11-dihydrocyclonerotriol, catenioblin C and sohirnone A (Fig. 6), which have been demonstrated to exhibit antifungal activity against Magnaporthe oryzae and C. albicans27.
CONCLUSION
The use of microbial antagonists for the biological management of plant pathogenic diseases is considered a potential alternative method. Potential fungal biocontrol agents against plant diseases include those belonging to the genus Trichoderma. There are hundreds of secondary metabolites known to be produced by useful fungi. Biocontrol fungi, including Trichoderma spp., secrete compounds that inhibit plant pathogens and are known to have a wide range of activities, including direct and indirect toxicity against plant diseases, activation of plant defences and stimulation of plant growth. Thus, treatment with Trichoderma metabolites considerably modifies the plant’s expressome, transcriptome and metabolic activity by altering particular pathways involved in the generation of important hormones, biotic/abiotic stress resistance and nutrient uptake. In this study, we concentrated primarily on the compounds that were produced by Trichoderma longibrachiatum and that were involved in interactions with phytopathogenic agents that resulted in advantageous outcomes for agricultural production. An intriguing alternative to the use of insecticides could be the application of certain metabolites with the aim of inducing host resistance and/or improving crop productivity.
SIGNIFICANCE STATEMENT
Among different biocontrol agents, Trichoderma species is well-studied and known for their ability to produce bioactive secondary metabolites such as polyketides, alkaloids, terpenoids and peptaibols against plant disease management. The secondary metabolites produced by Trichoderma have increased wide-ranging attention since they possess attractive chemical structures with notable biological activities. Certain species of the fungal genus Trichoderma are mycoparasites amongst them T. longibrachiatum has also been explored for use in combating fungal diseases on crops. The purpose of this review paper is to
succinctly review recent progress in the biopotential compounds of T. longibrachiatum and to provide a comprehensive overview with the secondary metabolites produced by T. longibrachiatum with emphasis on their chemistry and various bioactivities.
REFERENCES
- Mukherjee, P.K., B.A. Horwitz and C.M. Kenerley, 2012. Secondary metabolism in Trichoderma-a genomic perspective. Microbiology, 158: 35-45.
- Renshaw, J.C., G.D. Robson, A.P.J. Trinci, M.G. Wiebe, F.R. Livens, D. Collison and R.J. Taylor, 2002. Fungal siderophores: Structures, functions and applications. Mycol. Res., 106: 1123-1142.
- Karlsson, M., L. Atanasova, D.F. Jensen and S. Zeilinger, 2017. Necrotrophic mycoparasites and their genomes. Microbiol. Spectr., 5.
- O’Brien, P.A., 2017. Biological control of plant diseases. Australas. Plant Pathol., 46: 293-304.
- Fischer, G.J. and N.P. Keller, 2016. Production of cross-kingdom oxylipins by pathogenic fungi: An update on their role in development and pathogenicity. J. Microbiol., 54: 254-264.
- Vinale, F., K. Sivasithamparam, E.L. Ghisalberti, R. Marra and M.J. Barbetti et al., 2008. A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol. Mol. Plant Pathol., 72: 80-86.
- Viterbo, A., J. Invar, I. Hadar and I. Chet, 2007. Plant Disease Biocontrol and Induced Resistance via Fungal Mycoparasites. In: Environmental and Microbial Relationships (The Mycota, Volume 4), Kubicek, C.P. and I.S. Druzhinina (Eds.), Springer-Verlag, Berlin, Germany, ISBN: 978-3-540-71840-6, pp: 127-146.
- Daoubi, M., C. Pinedo-Rivilla, M.B. Rubio, R. Hermosa, E. Monte, J. Aleu and I.G. Collado, 2009. Hemisynthesis and absolute configuration of novel 6-pentyl-2H-pyran-2-one derivatives from Trichoderma spp. Tetrahedron, 65: 4834-4840.
- Chandra, P., R.K. Sharma and D.S. Arora, 2020. Antioxidant compounds from microbial sources: A review. Food Res. Int., 129: 108849.
- Demain, A.L., 2014. Importance of microbial natural products and the need to revitalize their discovery. J. Ind. Microbiol. Biotechnol., 41: 185-201.
- Khan, R.A.A., S. Najeeb, S. Hussain, B. Xie and Y. Li, 2020. Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic fungi. Microorganisms, 8: 817.
- Ming, Q., T. Han, W. Li, Q. Zhang and H. Zhang et al., 2012. Tanshinone IIA and tanshinone I production by Trichoderma atroviride D16, an endophytic fungus in Salvia miltiorrhiza. Phytomedicine, 19: 330-333.
- Vizcaíno, J.A., L. Sanz, R.E. Cardoza, E. Monte and S. Gutiérrez, 2005. Detection of putative peptide synthetase genes in Trichoderma species: Application of this method to the cloning of a gene from T. harzianum CECT 2413. FEMS Microbiol. Lett., 244: 139-148.
- Vinale, F., K. Sivasithamparam, E.L. Ghisalberti, S.L. Woo and M. Nigro et al., 2014. Trichoderma secondary metabolites active on plants and fungal pathogens. Open Mycol. J., 8: 127-139.
- Zin, N.A. and N.A. Badaluddin, 2020. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci., 65: 168-178.
- Al-Askar, A.A., E.M. Rashad, Z. Moussa, K.M. Ghoneem and A.A. Mostafa et al., 2022. A novel endophytic Trichoderma longibrachiatum WKA55 with biologically active metabolites for promoting germination and reducing mycotoxinogenic fungi of peanut. Front. Microbiol., 13: 772417.
- Du, F.Y., G.L. Ju, L. Xiao, Y.M. Zhou and X. Wu, 2020. Sesquiterpenes and cyclodepsipeptides from marine-derived fungus Trichoderma longibrachiatum and their antagonistic activities against soil-borne pathogens. Mar. Drugs, 18: 165.
- Zou, J.X., Y.P. Song and N.Y. Ji, 2021. Deoxytrichodermaerin, a harziane lactone from the marine algicolous fungus Trichoderma longibrachiatum A-WH-20-2. Nat. Prod. Res., 35: 216-221.
- Shi, W.L., X.L. Chen, L.X. Wang, Z.T. Gong and S. Li et al., 2016. Cellular and molecular insight into the inhibition of primary root growth of Arabidopsis induced by peptaibols, a class of linear peptide antibiotics mainly produced by Trichoderma spp. J. Exp. Bot., 67: 2191-2205.
- Kontani, M., Y. Sakagami and S. Marumo, 1994. First β-1,6-glucan biosynthesis inhibitor, bisvertinolone isolated from fungus, Acremonium strictum and its absolute stereochemistry. Tetrahedron Lett., 35: 2577-2580.
- Hewedy, O.A., K.S.A. Lateif, M.F. Seleiman, A. Shami, F.M. Albarakaty and R.M. El-Meihy, 2020. Phylogenetic diversity of Trichoderma strains and their antagonistic potential against soil-borne pathogens under stress conditions. Biology, 9: 189.
- Degani, O. and S. Dor, 2021. Trichoderma biological control to protect sensitive maize hybrids against late wilt disease in the field. J. Fungi, 7: 315.
- Sarsaiya, S., A. Jain, X. Fan, Q. Jia and Q. Xu et al., 2020. New insights into detection of a dendrobine compound from a novel endophytic Trichoderma longibrachiatum strain and its toxicity against phytopathogenic bacteria. Front. Microbiol., 11: 00337.
- Sridharan, A.P., S. Thangappan, G. Karthikeyan, S. Nakkeeran and S. Uthandi, 2021. Metabolites of Trichoderma longibrachiatum EF5 inhibits soil borne pathogen, Macrophomina phaseolina by triggering amino sugar metabolism. Microb. Pathogen., 150 :104714.
- Zhang, S., Y. Gan, J. Liu, J. Zhou and B. Xu, 2020. Optimization of the fermentation media and parameters for the bio-control potential of Trichoderma longibrachiatum T6 against nematodes. Front. Microbiol., 11: 574601.
- Zhang, S.H., J. Yang, H. Ma, Y. Yang and G.F. Zhou et al., 2021. Longibramides A-E, peptaibols isolated from a mushroom derived fungus Trichoderma longibrachiatum Rifai DMG-3-1-1. Chem. Biodivers., 18: e2100128.
- Sridharan, A.P., S. Thankappan, G. Karthikeyan and S. Uthandi, 2020. Comprehensive profiling of the VOCs of Trichoderma longibrachiatum EF5 while interacting with Sclerotium rolfsii and Macrophomina phaseolina. Microbiol. Res., 236: 126436.
- Xuan, Q.C., R. Huang, Y.W. Chen, C.P. Miao, K.X. Ma, T. Wang and S.H. Wu, 2014. Cyclonerol derivatives from Trichoderma longibrachiatum YM311505. Nat. Prod. Commun., 9: 313-314.
- Zhang, S., B. Xu, J. Zhang and Y. Gan, 2018. Identification of the antifungal activity of Trichoderma longibrachiatum T6 and assessment of bioactive substances in controlling phytopathgens. Pestic. Biochem. Physiol., 147: 59-66.
- Sallam, N.M.A., A.M.I. Eraky and A. Sallam, 2019. Effect of Trichoderma spp. on Fusarium wilt disease of tomato. Mol. Biol. Rep., 46: 4463-4470.
- Abo-Elyousr, K.A.M., S.I.I. Abdel-Hafez and I.R. Abdel-Rahim, 2014. Isolation of Trichoderma and evaluation of their antagonistic potential against Alternaria porri. J. Phytopathol., 162: 567-574.
- Rajani, P., C. Rajasekaran, M.M. Vasanthakumari, S.B. Olsson, G. Ravikanth and R.U. Shaanker, 2021. Inhibition of plant pathogenic fungi by endophytic Trichoderma spp. through mycoparasitism and volatile organic compounds. Microbiol. Res., 242: 126595.
- Lee, S., M. Yap, G. Behringer, R. Hung and J.W. Bennett, 2016. Volatile organic compounds emitted by Trichoderma species mediate plant growth. Fungal Biol. Biotechnol., 3.
- Gardiner, D.M., P. Waring and B.J. Howlett, 2005. The epipolythiodioxopiperazine (ETP) class of fungal toxins: Distribution mode of action functions and biosynthesis. Microbiology, 151: 1021-1032.
- Howell, C.R., R.D. Stipanovic and R.D. Lumsden, 1993. Antibiotic production by strains of Gliocladium virens and its relation to the biocontrol of cotton seedling diseases. Biocontrol Sci. Technol., 3: 435-441.
- Korpi, A., J. Järnberg and A.L. Pasanen, 2009. Microbial volatile organic compounds. Crit. Rev. Toxicol., 39: 139-193.
- Tarus, P.K., C.C. Lang’at-Thoruwa, A.W. Wanyonyi and S.C. Chhabra, 2003. Bioactive metabolites from Trichoderma harzianum and Trichoderma longibrachiatum. Bull. Chem. Soc. Ethiop., 17: 185-190.
- Serrano-Carreon, L., Y. Hathout, M. Bensoussan and J.M. Belin, 1992. Production of 6-pentyl-α-pyrone by Trichoderma harzianum from 18:n fatty acid methyl esters. Biotechnol. Lett., 14: 1019-1024.
- Andrade, R., W.A. Ayer and P.P. Mebe, 1992. The metabolites of Trichoderma longibrachiatum. Part 1. Isolation of the metabolites and the structure of trichodimerol. Can. J. Chem., 70: 2526-2535.
- Meng, J., W. Cheng, H. Heydari, B. Wang, K. Zhu, B. Konuklugil and W. Lin, 2018. Sorbicillinoid-based metabolites from a sponge-derived fungus Trichoderma saturnisporum. Mar. Drugs, 16: 226.
- Saif Ur Rehman, L.J. Yang, Y.H. Zhang, J.S. Wu and T. Shi et al., 2020. Sorbicillinoid derivatives from sponge-derived fungus Trichoderma reesei (HN-2016-018). Front. Microbiol., 11: 01334.
- Harned, A.M. and K.A. Volp, 2011. The sorbicillinoid family of natural products: Isolation, biosynthesis, and synthetic studies. Nat. Prod. Rep., 28: 1790-1810.
- Peng, J., X. Zhang, L. Du, W. Wang, T. Zhu, Q. Gu and D. Li, 2014. Sorbicatechols A and B, antiviral sorbicillinoids from the marine-derived fungus Penicillium chrysogenum PJX-17. J. Nat. Prod., 77: 424-428.
- Guo, W., J. Peng, T. Zhu, Q. Gu, R.A. Keyzers and D. Li, 2013. Sorbicillamines A-E, nitrogen-containing sorbicillinoids from the deep-sea-derived fungus Penicillium sp. F23-2. J. Nat. Prod., 76: 2106-2112.
- Abe, N., T. Arakawa and A. Hirota, 2002. The biosynthesis of bisvertinolone: Evidence for oxosorbicillinol as a direct precursor. Chem. Commun., 2002: 204-205.
- Ngo, M.T., M. van Nguyen, J.W. Han, M.S. Park, H. Kim and G.J. Choi, 2021. In vitro and in vivo antifungal activity of sorbicillinoids produced by Trichoderma longibrachiatum. J. Fungi, 7: 428.
How to Cite this paper?
APA-7 Style
Ramasamy,
P., Subbu,
B., Narayanasamy,
P. (2023). Trichoderma longibrachiatum’s Secondary Metabolite-A Review on Potential Compounds for Plant Growth and Biological Control for Plant Pathogens. Asian Journal of Biological Sciences, 16(3), 202-211. https://doi.org/10.3923/ajbs.2023.202.211
ACS Style
Ramasamy,
P.; Subbu,
B.; Narayanasamy,
P. Trichoderma longibrachiatum’s Secondary Metabolite-A Review on Potential Compounds for Plant Growth and Biological Control for Plant Pathogens. Asian J. Biol. Sci 2023, 16, 202-211. https://doi.org/10.3923/ajbs.2023.202.211
AMA Style
Ramasamy
P, Subbu
B, Narayanasamy
P. Trichoderma longibrachiatum’s Secondary Metabolite-A Review on Potential Compounds for Plant Growth and Biological Control for Plant Pathogens. Asian Journal of Biological Sciences. 2023; 16(3): 202-211. https://doi.org/10.3923/ajbs.2023.202.211
Chicago/Turabian Style
Ramasamy, Priya, Balachandar Subbu, and Prabhakaran Narayanasamy.
2023. "Trichoderma longibrachiatum’s Secondary Metabolite-A Review on Potential Compounds for Plant Growth and Biological Control for Plant Pathogens" Asian Journal of Biological Sciences 16, no. 3: 202-211. https://doi.org/10.3923/ajbs.2023.202.211

This work is licensed under a Creative Commons Attribution 4.0 International License.